#Tálamo Óptico
Text
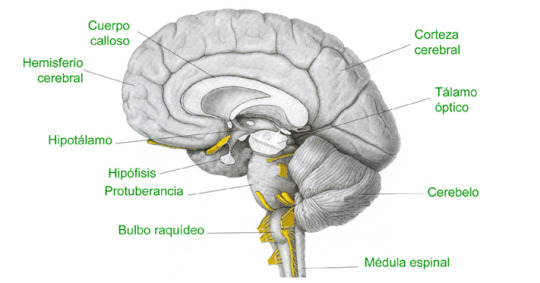
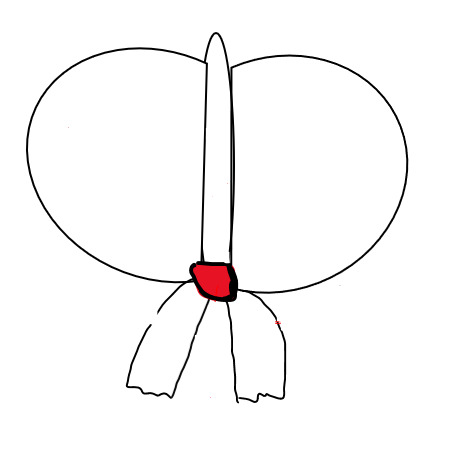
#Cerebro#Brain#Cuerpo Calloso#Hard Body#Hemisferio Cerebral#Cerebral Hemisphere#Hipotálamo#Hypothalamus#Hipófisis#Hypophysis#Protuberancia#Protuberance#Bulbo Raquídeo#Medulla Oblongata#Corteza Cerebral#Cerebral Cortex#Tálamo Óptico#Optic Thalamus#Cerebelo#Cerebellum#Médula espinal#Spinal Cord
0 notes
Text
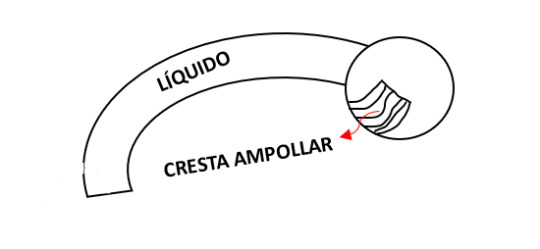
PLEXOS COROIDEOS Y LÍQUIDO CEFALORRAQUÍDEO
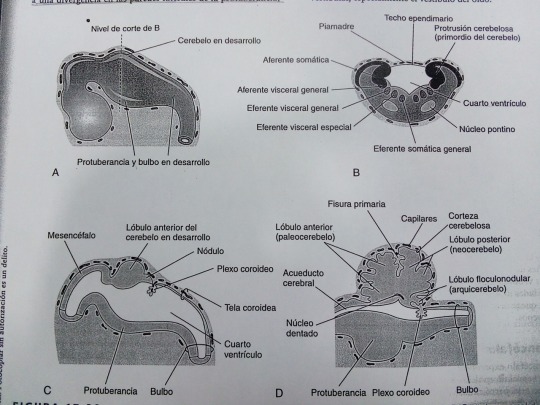
El fino techo ependimario del 4to ventrículo esta cubierto externamente de por la piamadre, derivada del mesénquima asociado al rombencéfalo. Esta membrana vascular junto con el techo ependimario forman la tela coroidea (recubrimiento del 4to ventrículo).
Los plexos coroideos segregan líquido ventricular que se convierte en líquido cefalorraquídeo (LCR).
En el LCR y los plexos coroideos se encuentra diferentes morfogenos señalizadores importantes para el desarrollo del encéfalo.
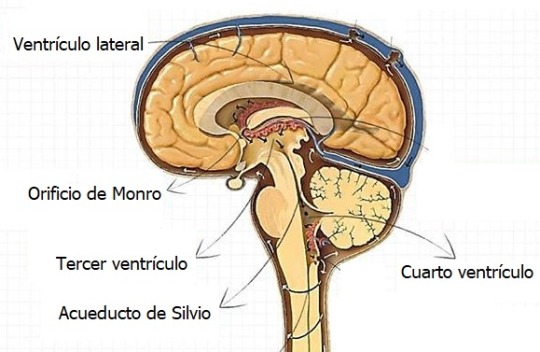
El techo del 4to ventrículo presenta evaginaciones que se rompen para formar los:
Agujeros de Magendie y Luschka (aberturas medial y lateral). Los cuales permiten la entrada del LCR procedente del 4to ventrículo en el espacio subaracnoideo.
La principal zona de absorción del LCR hacia el sistema venoso la constituyen las vellosidades aracnoideas (protrusiones en los senos venosos durales).
MESENCÉFALO
Experimenta menos cambios que cualquier otra parte del encéfalo en desarrollo,execto en lo que se refiere a la parte caudal del rombencéfalo.
El canal neural presenta un estrechamiento originándose el acueducto cerebral que pone en conexión el 3er y 4to ventrículo.
Los Neuroblastos migran desde las placas alares del mesencéfalo hasta el techo acumulan formando los colículos superiores e inferiores bilaterales (implicados en los reflejos visuales y auditivos). Los neuroblastos de las placas basales dan lugar a grupos de neuronas en el tegmento del mesencéfalo.
Las fibras que crecen a partir del cerebro forman los pedúnculos cerebrales en la parte anterior. Los cuales aumentan progresivamente su tamaño medida que incrementa el número de fibras descendentes que atraviesan el mesencéfalo en desarrollo hasta el tronco encefálico y la medula espinal.
PROSENCÉFALO
A medida que se cierra en el neuroporo rostral aparecen la vesículas ópticas a cada lado del prosencéfalo.
⇨ Primordios de la retina y los nervios ópticos
Tiempo después aparecen el segundo par de divertículos vesículas telencefálicas.
⇨ Primordios de los hemisferios cerebrales y sus cavidades se convierten en ventrículos laterales
Las cavidades del telencéfalo y el diencéfalo contribuyen a la formación del 3er ventrículo.
DIENCÉFALO
En las paredes laterales del tercer ventrículo aparecen 3 protrusiones:
• Tálamo
• Hipotálamo
• Epitálamo
El tálamo se desarrolla rápidamente a cada lado del 3er ventrículo. Y se encuentran y fusionan en la línea media en el 70% de los encéfalos formando la comisura intertalámica a través del 3er ventrículo (puente ausente en el 20% de los encéfalo).
0 notes
Text
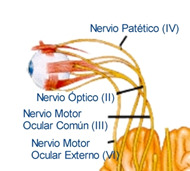
Óptico (II)
Función: Enteramente sensitivo, sus función es la visión.
Origen: Proviene de las células ganglionares de la retina y termina en los cuerpos geniculados del tálamo.
3 notes
·
View notes
Text
Morfologia III
Neurônios: Células mais importantes no tecido nervoso, capazes de receber, estimular, além de produzir respostas no sistema nervoso. É formado por pericário (onde se forma o núcleo e é mantido o material genético) e prolongamentos celulares que podem ser dendritos - prolongamentos celulares múltiplos que contribuem para o aumento da superfície receptora dos neurônios e que recebem as informações
ou axônios - conduzem os impulsos nervosos do pericário até ao ponto em que serão emitidos para outras células, os neurônios podem ser unipolares, bipolares e multipolares
Transporte axônio: Transporte de substâncias ao longo dos prolongamentos (do pericário até as terminações axônicas) influenciados pelos microtúbulos e neurofilamentos
No pericário ocorrem as atividades metabólicas essenciais para a vida das células, os prolongamentos que são separados das células acabam se degenerando e morrendo
Oligodendrócitos: Responsável pela formação da bainha de mielina no SNC
Células de Schwann ou neurolemócitos: Faz a mielinização no SNP
Sinapse: Faz com que as células nervosas entrem em contato e se comuniquem com outras células, quando um impulso nervoso chega a uma terminação nervosa ocorre a liberação de um neurotransmissor que age sobre os receptores da membrana pós-sináptica
As sinapses podem ser excitatórias ou inibitórias
Neuróglias ou células de glia: Células que não estão envolvidas na recepção e condução de impulsos nervosos. Pode ser astróglia (ajuda no processo de cicatrização do SNC, nutrição e manutenção do tecido), oligodendróglia (produção de mielina para os axônios), micróglia (removem detritos e microrganismos do SNC) e epêndima (reveste as cavidades do SNC).
Nervos: Constituídos por fibras nervosas (axônios), possuem bainha de mielina
Terminações nervosas: Sensíveis a determinados tipos de estímulo que irão determinar reações nervosas nas fibras do SNC
Córtex cerebral: Região rica em neurônios, a espécie humana é privilegiada com uma grande quantidade de córtex cerebral que os possibilitam de desenvolver algumas funções como fala, raciocínio e planejamento que outras espécies não conseguem.
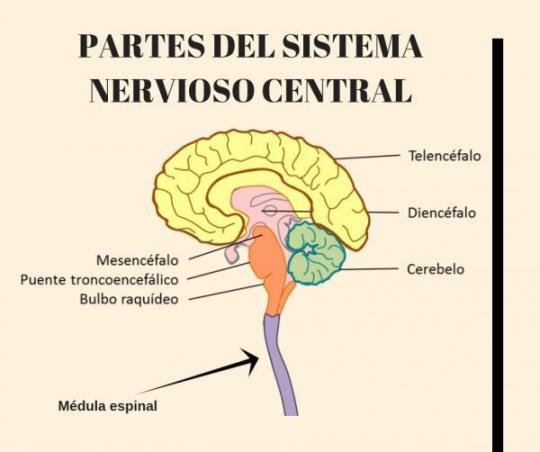
Sistema nervoso central: Encéfalo e medula espinhal. O encéfalo é dividido em três estruturas chamadas cérebro que é dividido em telencéfalo e diencéfalo, tronco encefálico que é dividido em ponte, bulbo e mesencéfalo, e cerebelo que tem função de promover controle de motricidade (dividido em cerebelo espinhal que promove manutenção do tônus para manter a postura corporal, vestibular que promove equilíbrio e cortical que é importante no planejamento de motricidade e coordenação)
Sistema nervoso periférico: Se comunica com o SNC através dos nervos (tem origem na medula espinhal ou no encéfalo) e gânglios
Sistema nervoso: Nos vertebrados tem origem no folheto embrionário, no ectoderma.
Meninges: Membranas conjuntivas, presente na medula espinhal
Fascículos grácil que tem função de movimentar os membros inferiores e fascículo cuneiforme que movimenta os membros superiores
Diencéfalo: Origina o tálamo - corpo geniculado lateral (visão), corpo geniculado medial (audição), aderência intertalâmica
subtálamo - Núcleos da base do cérebro são massas cinzentas, corpo estriado (faz conexão com o giro pré central, controle motor), putâmen, corpo amigdaloíde (associado ao controle de emoções)
epitálamo - Habênulas, corpo pineal (ritmo circadiano, influência o metabolismo)
hipotálamo - Quiasma óptico, corpos mamilares
Lobos do cérebro: Frontal, parietal (giro pós central tbm conhecido como área de broadman 1 2 3 com função somatossensorial, giro pré central com função motora área 4), temporal (giro temporal transverso função auditiva, área 41 e 42 de brodmann) e ocipital
Irrigação encefálica: Artéria cerebral anterior e média (originárias da artérias carótidas internas), a. cerebral posterior, a. carótida interna direita e esquerda, a. basilar (irriga tronco encefálico e dá origem a a. cerebral posterior), a. cerebelar, a. vertebral direita e esquerda (irriga a medula e cerebelo e dá origem a a. basilar)
Substância negra: Mesencéfalo, produz dopamina no cérebro.
Colículo superior: Junto com o corpo geniculado lateral tem função de estimular a visão
Colículo inferior: Junto com o corpo geniculado medial tem função de estimular a audição
Núcleo basal de meynert: Situado na substância inominata, a degeneração dos núcleos está associado a doença de Alzheimer
Classificação de Brodmann: Divide o córtex em 52 regiões designadas por números que ajudam a separar as áreas por funções distintas,
2 notes
·
View notes
Text
Pretectum (región cerebral): ubicación, funciones y patologías
El [mesencéfalo](/neurociencias/mesencefalo) es una parte del cerebro imprescindible para la homeóstasis de nuestro organismo y para nuestra supervivencia. En su interior podemos localizar el pretectum, un área implicada en el procesamiento visual inconsciente y relacionada con procesos automáticos como el reflejo oculomotor o [el sueño REM](/neurociencias/fase-rem). **En este artículo te explicamos qué es el pretectum, dónde se ubica y cómo está estructurada esta región cerebral**. Además, se exponen las principales funciones que desempeña, y un ejemplo de uno de los trastornos más habituales tras la lesión de esta área del cerebro. ## Pretectum: definición, ubicación y estructura El área pretectal o pretectum es una región cerebral ubicada en el mesencéfalo, una estructura que une el tronco encefálico con el [diencéfalo](/neurociencias/diencefalo). Esta área forma parte del sistema visual subcortical y posee conexiones recíprocas con la retina. Está compuesto por varios núcleos altamente interconectados. En el mesencéfalo se localiza el tectum, una estructura situada en su parte posterior, compuesto por dos colículos superiores y dos inferiores. **El pretectum se localiza en la parte anterior del colículo superior y posterior al tálamo** (centro de relevo de la información sensorial que se dirige a la corteza cerebral), y sobre la sustancia gris periacueductal y el núcleo de la comisura posterior. Aunque no se han podido delimitar claramente, **los siete núcleos del pretectum reciben cada uno de ellos un nombre con su respectiva región**; los cinco núcleos primarios son: el núcleo pretectal olivar, el núcleo del tracto óptico, el núcleo anterior, el núcleo medial y el núcleo posterior. Además, se han identificado otros dos núcleos adicionales: el área comisural pretectal y los limitans posteriores. Aunque estos dos últimos núcleos no se han estudiado del mismo modo que los cinco núcleos primarios, las investigaciones realizadas han demostrado que los dos núcleos adicionales reciben conexiones de la retina, lo que sugiere que también jugarían un papel en el procesamiento de la información visual. Para aclarar esto, a continuación veremos qué funciones desempeñan los núcleos pretectales. ## Funciones **El pretectum forma parte del sistema visual subcortical y las neuronas de esta estructura responden a intensidades variables de iluminación**. Las células de los núcleos pretectales están implicadas, principalmente, en la mediación de respuestas conductuales inconscientes a los cambios agudos en la luz. En general, estas respuestas incluyen el inicio de determinados reflejos optocinéticos, aunque, como veremos más adelante, el pretectum también participa en otros procesos como la regulación de la nocicepción (la codificación y el procesamiento de los estímulos potencialmente dañinos o dolorosos) o el sueño REM. ### 1. El reflejo fotomotor **El reflejo fotomotor o reflejo pupilar de la luz se produce cuando la pupila de ojo responde a los estímulos lumínicos**, aumentando o disminuyendo su diámetro. Este reflejo está mediado por varios de los núcleos del pretectum, en particular el núcleo pretectal olivar, que reciben información del nivel de luz de la retina ipsilateral a través del tracto óptico. Los núcleos pretectales aumentan gradualmente su activación en respuesta a los niveles crecientes de iluminación, y esta información se transmite directamente al núcleo de Edinger-Westphal, que se encarga de transmitir los impulsos nerviosos y de mandar la señal al esfínter pupilar, a través del ganglio ciliar, para que se produzca la contracción pupilar. ### 2. Movimientos oculares de seguimiento Los núcleos pretectales, y en particular el núcleo del tracto óptico, están involucrados en la coordinación de los movimientos oculares durante el seguimiento ocular lento. Estos movimientos permiten que el ojo siga de cerca un objeto en movimiento y lo alcance después de un cambio inesperado en la dirección o la velocidad. **Las neuronas de la retina que son sensibles a la dirección y se ubican en el núcleo del tracto óptico**, proporcionan información de los errores de desplazamiento horizontal en la retina a través de la oliva inferior. Con luz del día, esta información se detecta y se transmite por neuronas que poseen grandes campos receptivos, mientras que las neuronas parafoveales con pequeños campos receptivos lo hacen en cuando hay oscuridad o en penumbra. Es de este modo mediante el cual el núcleo del tracto óptico es capaz de enviar la información de los errores en la retina para guiar los movimientos oculares. Además de su papel en el mantenimiento de dichos movimientos oculares, el pretectum se activa durante el nistagmo optocinético en el que el ojo vuelve a una posición central, orientada hacia adelante, después de que un objeto que estaba siendo seguido se salga del campo de visión. ### 3. Antinocicepción **El núcleo pretectal anterior participa en la disminución activa de la percepción de los estímulos dolorosos o antinocicepción**. Aunque el mecanismo por el cual el pretectum altera la respuesta del organismo a estos estímulos aún se desconoce, las investigaciones sugieren que la actividad del núcleo pretectal anterior ventral incluye a neuronas colinérgicas y serotoninérgicas. Estas neuronas activan vías descendentes que hacen sinapsis en la médula espinal e inhiben las células nociceptivas en su asta dorsal. Además de su mecanismo antinociceptivo directo, el núcleo pretectal anterior envía proyecciones a regiones del cerebro que, a través de conexiones de la corteza somatosensorial, regulan la percepción del dolor. Dos de estas regiones que se sabe que proyecta el pretectum son la zona incierta (un núcleo del subtálamo) y el núcleo posterior del tálamo. Diversos estudios han encontrado que el núcleo pretectal anterior dorsal disminuye de forma más significativa la percepción del dolor breve, mientras que su parte ventral lo haría en casos de dolor crónico. Debido a su papel en la reducción del dolor crónico, se ha sugerido que la actividad anormal de este núcleo pretectal podría estar implicada en el síndrome de dolor neuropático central. ### 4. Sueño REM En lo que respecta al sueño de movimientos oculares rápidos o sueño REM, **las investigaciones apuntan a que múltiples núcleos pretectales podrían estar involucrados en la regulación de este tipo de sueño** y de otros comportamientos similares. Se ha sugerido que el pretectum, junto con el colículo superior, puede ser responsable de provocar alteraciones no circadianas en las conductas relacionadas con el sueño REM. Los estudios en animales, concretamente en ratas albinas, han mostrado que los núcleos pretectales que reciben información de la retina, en particular el núcleo del tracto óptico y el núcleo pretectal posterior, son en parte responsables de iniciar el sueño de movimientos oculares rápidos. El descubrimiento de la existencia de proyecciones desde el pretectum a varios núcleos talámicos involucrados en la activación cortical durante el sueño REM, específicamente al núcleo supraquiasmático que es parte de un mecanismo regulador de este tipo de sueño, respaldaría esta última hipótesis. ## El síndrome pretectal El síndrome pretectal, también denominado síndrome de Parinaud o síndrome mesencefálico dorsal, hace referencia a un conjunto de signos y síntomas clínicos que incluyen alteraciones en la motilidad ocular y cuya principal causa es la afectación del pretectum y otras [áreas cerebrales](/neurociencias/partes-cerebro-humano) adyacentes. Este síndrome provoca alteraciones en la motilidad ocular, tanto a nivel externo como a nivel interno. Los signos más habituales son los siguientes: * **Alteraciones pupilares**: asimetría del tamaño de las pupilas, reacción disociada a la luz, paresia acomodatoria y arreflexia pupilar. * **Parálisis de la mirada vertical** hacia arriba (de tipo supranuclear). * **Retracción del párpado** (signo de Collier). * **Signo de lid lag**: cuando el párpado superior no puede mantener su posición relativa respecto al globo ocular (al mover los ojos hacia abajo). * **Nistagmo de convergencia-retracción**: cuando la persona intenta mirar hacia arriba, los ojos vuelven a su posición central y los globos oculares se retraen. #### Referencias bibliográficas: * Gamlin, P. D. (2006). The pretectum: connections and oculomotor-related roles. Progress in brain research, 151, 379-405. * Keane, J. R. (1990). The pretectal syndrome: 206 patients. Neurology, 40(4), 684-684. * Miller, A. M., Miller, R. B., Obermeyer, W. H., Behan, M., & Benca, R. M. (1999). The pretectum mediates rapid eye movement sleep regulation by light. Behavioral neuroscience, 113(4), 755.
Ver Fuente
Ver Fuente
0 notes
Video
youtube
La fisiología de la #visión más allá de la retina, es decir la anatomía del nervio óptico, el quiasma óptico y su relación con el hipotálamo y el ciclo circadiano. Hoy revisamos la fisiología de la visión más allá de la retina, es decir la anatomía del nervio óptico, el quiasma óptico y su relación con el hipotálamo y el ciclo circadiano; más adelante veremos la función del tálamo y la corteza cerebral, para finalmente revisar algunas patologías como hemianopsia, la ceguera cortical y el jet lag.
0 notes
Text
Bouquet & a Comunicação Olfatória

Bouquet é um projeto de interface olfatória sintética criado pelos estudantes Erika Marthins, Arthur Moscatelli, Pietro Alberti e Andrea Ramìrez Aburto da ÉCAL na Suíça, surgido de um workshop de exploração de comportamentos em comunicação com o objetivo de desenvolver dispositivos para comunicação além do smartphone. O projeto foi desenvolvido em apenas uma semana e visa criar uma relação sinestésica da interface com a obra.
Como?
A interface funciona através de um cone de madeira com um buraco na base e um sensor óptico na ponta, conectado a um Arduino detectando as cores e identificando os cheiros associados. Dentro do cone um disco controlado por um motor maneja os cheiros diretamente no nariz do usuário, rotacionando pedaços de algodão imbuídos de diferentes aromas de acordo com as cores detectadas. O cone é utilizado em conjunto com uma série de pôsteres de cores chapadas para serem traduzidas em cheiros.

Por que?
O projeto cria uma forma interação pouco explorada em projetos de design, aonde o aspecto visual fica em segundo plano para a comunicação olfatória. O olfato é um dos sentidos menos explorados em mídias, e também o sentido mais associado a memória e emoção humana, por isso a área que mais tem sido bastante explorado seria a de branding pela sua capacidade de influenciar consumidores a passarem mais tempo em lugares, mudar a perspectiva sobre produtos e criar conexões emocionais com marcas que ficam marcadas na memória, como uma forma de logo olfatório.
A relação entre cheiro e emoção pode ser entendida a partir da investigação do processamento das informações olfativas pelo sistema sensorial. Quando sentimos um aroma, de imediato as amígdalas trabalham e relacionam aquele odor à ação que está ocorrendo ou como nos sentimos naquele momento. (...) Quando voltamos a sentir o mesmo cheiro, a memória afetiva é ativada, e a conexão entre o aroma e a emoção correspondente torna-se perceptível.
- Scardua (2011, s.p)
Com diversos projetos focando no desenvolvimento de visuais realísticos e imersivos, e mais recentemente no feedback tátil, muito voltado para a integração em tecnologias de realidade aumentada e realidade virtual; o projeto aponta uma possibilidade de integração do olfato nessas tecnologias, aonde o olfato pode se integrar a narrativa de um jogo ou a interação de um software, integrando um outro nível de leitura da experiência, até mais rápida do que a visão ou audição.
O olfato é nosso sentido mais lento, porque depende de mensagens carregadas pelo ar, mas ele também pode ser nosso sentido mais rápido. (...) Os sinais visuais ou sonoros devem ser assimilados pelo tálamo antes de chegar às regiões interpretativa do cérebro, enquanto mensagens olfativas vão diretamente do nariz para o córtex olfativo do cérebro, para processamento instantâneo.
- Jay A. Gottfried
vimeo
Futuro
Como é um projeto feito em curto tempo não é muito polido e apesar da simplicidade tem um aspecto um pouco desajeitado na interação com o usuário, sendo bastante grande criando uma certa dificuldade de manejá-lo e tendo um formato não muito intuitivo, apesar de remeter ao nome e sua função como um buquê de aromas. Com um motor menor talvez poderia ter sido desenvolvido um recipiente mais enxuto e ergonômico para o manejo do indivíduo, com afinamento no meio do cone formando uma área mais confortável para as mãos. Ao invés de um pequeno buraco poderia ter sido usado algum tipo de tela que fornecesse vazão e mais proteção ao objeto, não possibilitando a entrada de insetos ou outras coisas indesejadas.
A solução do disco com essências artificiais diferentes é bastante efêmera e não seria uma solução eficaz para uma tecnologia de uso mais frequente. Também é muito provável dos cheiros se misturarem, visto que há pouca barreira entre eles, e até de serem absorvidos pelo cone por ser de madeira, que comumente absorve os cheiros do que é colocado no material, fazendo com que os componentes tenham de ser mudados de recipiente. Um recipiente menos absorvente, como algum tipo de metal poderia ajudar a manter os cheiros livres de influência por mais tempo. Além disso os discos poderiam ter formas mais compactadas, e o motor ser usado não só para rotacionar como abrir os receptáculos de cheiros, os mantendo mais frescos e livre de influência de outros cheiros contidos na interface. Os aromas poderiam estar guardados em cápsulas ao invés de recipientes de algodão, assim como o Scentee, e para aumentar as possibilidades de cheiros poderia ser usado a interação entre dois cheiros em cores no meio do caminho entre as definidas.
Se aplicado a tecnologia VR e AR a interação sinestésica com os cheiros e a maior possibilidade de tipos de cheiros e combinações poderia ser transformando em uma mecânica de jogabilidade principalmente guiada pelo olfato, e a interface poderia ser pensada para acoplar ao óculos e ao controle, utilizando o controle da mão para a detecção de cores; usando um celular para captar cores reais no caso do AR e no caso do VR apenas lendo o código das cores dentro do jogo, e acoplando a saída do cheiro ao óculos ou ao smartphone como faz o Scentee. Acoplado as tecnologias háptica de resposta de toque já em desenvolvimento para essas plataformas, 4 dos nossos 5 sentidos seriam utilizados nessas experiência, aumentando a capacidade imersiva da tecnologia.

Bibliografia:
Fast Company, Smell Based User Interfaces are on the way
Niklas Roy, ECAL Communication Devices Estadão, Olfato é o Sentido Mais Ligado as Emoções e a Memória
Memória Olfativa
0 notes
Photo
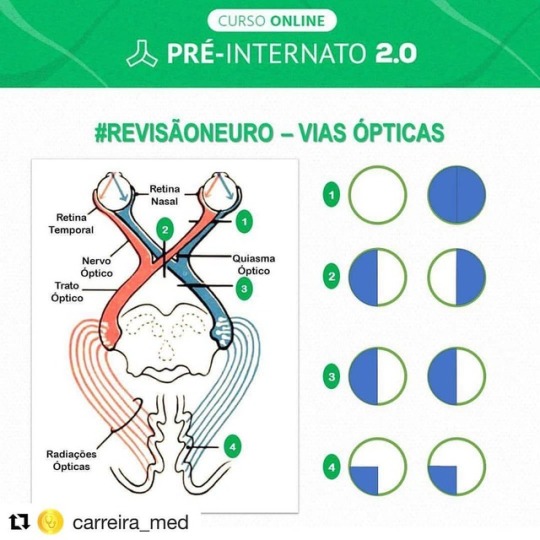
Visual Pathway #Repost @carreira_med A luz proveniente dos campos visuais incide na retina. A parte medial da retina é chamada de retina nasal, enquanto a parte lateral é chamada de retina temporal. Na retina nasal, a luz que chega é do campo temporal, e vice versa O impulso nervoso gerado segue através do Nervo óptico (NC II). No quiasma óptico, as fibras provenientes da retina nasal (campos temporais) irão se cruzar, e passar a fazer parte do trato óptico contralateral. Os tratos ópticos seguem até o teto do mesencéfalo, e de lá partem para os corpos geniculados laterais do tálamo. As fibras das radiações ópticas saem do tálamo em direção aos lábios da fissura calcarina, onde o impulso nervoso será interpretado! #Neuroanatomia #Medicina #ViasÓpticas #Neurologia #Neurocirurgia #Internato #neurosurgeryblog #neurocirurgiabr
#neurosurgeryblog#medicina#viasópticas#neurologia#internato#repost#neurocirurgia#neurocirurgiabr#neuroanatomia
0 notes
Text
Sin la corteza visual de su cerebro un Niño puede ver
Nuevo artículo publicado en https://www.prozesa.com/2017/12/10/nino-puede-ver-sin-la-corteza-visual-de-su-cerebro/
Sin la corteza visual de su cerebro un Niño puede ver
Un niño de siete años ha sorprendido a la medicina. Había perdido la corteza visual del cerebro, lo que suponía que debía haberse quedado ciego. Sin embargo, el chico puede ver lo suficientemente bien como para reconocer rostros y moverse con soltura. La historia la han contado esta semana los investigadores del Instituto de Medicina Regenerativa de Australia en la Universidad de Monash durante una conferencia de neurociencia en Sydney. Al Parecer, BI (las iniciales del niño) sufrió una lesión bilateral en el lóbulo occipital cuando tenía tan solo dos semanas de edad. Por tanto, tenía un daño extremadamente grave en su corteza visual. Esta es una región de la parte externa del cerebro situada en la zona posterior de la cabeza, la cual es responsable de clasificar la información de la retina en imágenes personales. La retina envía mensajes por el nervio óptico a un centro de transmisión en el tálamo llamado pulvinar inferior, que ayuda a controlar y enfocar los ojos en función de los objetos en la línea de visión. Desde ahí, la información viaja a una parte de la corteza visual denominada como área visual 1 (V1). Ocurre que cuando BI era un bebé sufrió una deficiencia de acil-CoA deshidrogenasa de cadena media (MCAD), una condición o trastorno genético que impide que los tejidos conviertan ciertas grasas en energía. Como resultado de ello, le dejaron una lesión en su V1, lo que quiere decir que debería haberse quedado ciego, o como mínimo con la incapacidad de convertir la información retinal en una imagen coherente. Pero parece que no. Según explica el neurocientífico Iñaki-Carril Mundiñano: Anteriormente, se sabía que las lesiones en V1 podían dejar en los afectados ciertos rasgos visuales conocidos como “visión ciega”. Algo así como “ver sin ser consciente de ninguna imagen”. En esos casos, se presenta como una habilidad para responder a estímulos visuales o cosas “sensoriales” como el movimiento. Lo extraordinario de la “visión ciega” es que demuestra que nuestro sentido de la vista se extiende mucho más allá de lo que experimentamos conscientemente como imágenes. No en el caso de BI, quién experimenta imágenes reales dentro de su mente. Para Mundiñano: Aunque ahora mismo todo son hipótesis, los investigadores apuestan por una idea: es posible que como la lesión se produjo cuando BI era muy joven, su cerebro en desarrollo fuera capaz de reencaminar la información de la retina para hacer uso de una de las otras unidades de la corteza visual. Las primeras pruebas a través de imágenes de resonancia magnética (MRI) han mostrados resultados satisfactorios sobre esta hipótesis. De ser así, el cerebro y el cuerpo humano son todavía más asombrosos de lo que ya pensábamos.
0 notes
Text
Neurología animal
Neurología. Sistema nervioso. La neurona
Neurología es la parte de la Anatomía Sistemática que se ocupa del sistema nervioso.
El sistema nervioso está formado por los órganos encargados de regir todos los procesos vitales del ser vivo, relacionando armónicamente cada uno de los órganos. Para ello, une y coordina la acción de diferentes partes corporales, capta estímulos, tanto del exterior como del interior, emite respuestas rápidas ante estos estímulos las cuales pueden ser de índole motora, secretora, sensitiva e incluso síquica y de este modo realiza la integración del organismo.
El sistema nervioso puede dividirse desde el punto de vista morfológico en dos porciones: un Sistema Nervioso Central en el interior de una protección ósea formada por el canal vertebral y la cavidad craneana y un Sistema Nervioso Periférico por fuera de dicha protección. Al sistema nervioso central corresponde el encéfalo y la médula espinal y al sistema nervioso periférico los ganglios y los nervios.
Esta división del sistema nervioso, sin embargo, no está acorde con un criterio funcional, de ahí que desde este punto de vista sea más adecuado considerar un sistema nervioso somático o sistema nervioso de la vida de relación, voluntario y encargado de contactar con el medio exterior, comunica al animal con el medio ambiente, interviene en la locomoción, sus procesos son fundamentalmente voluntarios, y otro sistema nervioso de la vida vegetativa o autónomo, en gran medida independiente de la voluntad, capaz de regular la actividad interna del ser vivo, interviene en la regulación de las estructuras viscerales que son involuntarias y automáticas, controla los procesos de secreción, digestión, excreción, contracción de músculos lisos, reproducción, actividad cardiaca, glandular, etc.
El sistema nervioso funciona basado en impulsos eléctricos, interviene en todos los procesos fisiológicos obrando como mediador entre el estímulo y la respuesta.
Esta actividad tiene tres fases:
1. Recepción de los estímulos
2. Transformación, conducción y elaboración de los mismos
3. Transmisión de los impulsos (respuesta) a los órganos efectores
Los receptores son órganos especializados en captar los estímulos, estos estímulos viajan por vías aferentes hacia el SNC donde se decodifican y coordinan adecuadamente y luego éste envía una respuesta por sus vías eferentes hacia los órganos efectores que realizarán las acciones que le son concernientes.
La célula nerviosa se llama neurona, consta de un cuerpo y prolongaciones: axón y dendritas. Las dendritas conducen impulsos hacia el cuerpo celular (centrípetos) y el axón lleva impulsos desde el cuerpo celular hacia el exterior (centrífugos). Cada neurona posee un solo axón y puede tener una o varias dendritas. Las dendritas son prolongaciones cortas, en cambio el axón por lo general es muy largo. Los nervios están formados por axones de muchas neuronas envueltos en una vaina llamada epineuro.
Los estímulos viajan por las prolongaciones por medio de impulsos eléctricos pero las uniones entre prolongaciones o entre éstas y un órgano receptor o efector se realiza por medio de un mecanismo llamado sinapsis por medio del cual se opera el influjo nervioso de una neurona a otra. Esta sinapsis se produce debido a que las ramificaciones terminales de la prolongación nerviosa posee unos bulbos sinápticos o botones terminales los cuales poseen numerosas vesículas que contienen sustancias neurotransmisoras que son liberadas por el impulso nervioso y cuya liberación permite el paso del flujo nervioso.
Sistema nervioso central – Medula Espinal – Encéfalo - Meninges
Está formado por el encéfalo (parte del SNC encerrado en la cavidad craneana) y la médula espinal (parte del SNC encerrada en el canal medular de la columna vertebral), ambos están envueltos por las meninges.
Los grupos de cuerpos neuronales se distinguen por su color más oscuro, sobre todo si se los contrasta con la blancura de los haces de fibras adyacentes; esto permite distinguir entre la sustancia gris y blanca del SNC.
En el encéfalo la sustancia gris se encuentra en la periferia y la blanca en posición central, mientras que en la médula espinal la sustancia gris ocupa una posición central con respecto a la blanca.
Las aglomeraciones aisladas de cuerpos neuronales dentro del encéfalo se conocen como núcleos.
Los haces de fibras de origen destino y función común se congregan dentro del encéfalo y médula en fascículos o tractos, aunque normalmente los límites de éstos no son evidentes y sólo se pueden establecer con medios experimentales. La mayoría de estos tractos se designan por la combinación de su origen que se emplea como prefijo y su destino que se emplea como sufijo, en consecuencia inmediatamente se sabe el significado de nombres como tractos espinocerebeloso y cerebroespinal.
El encéfalo está alojado en el neurocráneo y su forma refleja en general a la cavidad craneana. Las divisiones del mismo son:
PROSENCÉFALO
🐎Telencéfalo
🐎Palium
🐎Rinencéfalo
🐎Cuerpo estriado
🐎Diencéfalo
🐎Epitálamo
🐎Tálamo
🐎Hipotálamo
MESENCÉFALO
🐇Mesencéfalo
🐇Tubérculos cuadrigéminos
🐇Pedúnculos cerebrales
ROBENCÉFALO
🐭Metencéfalo
🐭Puente
🐭Cerebelo
🐭Mielencéfalo
🐭Médula oblonga
La médula espinal se encuentra en el canal vertebral. Es un cordón cilíndrico un poco aplanado dorso ventralmente que se extiende desde el agujero magno hasta la mitad del sacro. Al corte transversal presenta una porción periférica de sustancia blanca y una central de sustancia gris, dividida a su vez en una porción dorsal sensitiva y una ventral motora.
Las meninges son las tres membranas que envuelven y protegen al encéfalo y a la médula espinal. Sus nombres son desde afuera hacia adentro duramadre, aracnoides y piamadre. La primera es más gruesa y fibrosa y las dos últimas son más delgadas y vascularizadas. La piamadre espinal está muy unida a la médula y la piamadre craneal lo está al encéfalo. La función de las meninges es proteger al encéfalo y a la médula espinal.
Sistema nervioso periférico
Consta de un número variable de nervios espinales según la especie y doce nervios craneales. Está constituido por fibras nerviosas y ganglios nerviosos, estos últimos son aglomeraciones de cuerpos neuronales en nervios periféricos y pueden formar engrosamientos visibles que se distinguen por su color y textura, más oscuros y firmes que la de los respectivos troncos nerviosos.
Formación y distribución de los nervios espinales:
De la sustancia gris de la médula espinal parten dos raíces, una ventral motora y una dorsal sensitiva. Las fibras de la raíz dorsal convergen para formar el ganglio espinal que es externo respecto de la duramadre y se halla en el agujero intervertebral o próximo a él.
Más allá del ganglio las raíces dorsal y ventral se unen para formar el tronco del nervio espinal que emerge por el agujero intervertebral. Cada nervio en el punto de emergencia emite una pequeña rama meníngea que se une a fibras procedentes del ramo comunicante (ver definición abajo) y penetra en el canal vertebral en el cual se distribuye.
El nervio luego se divide en una rama dorsal y otra ventral. Por lo general las ramas ventrales son más voluminosas, excepto en la región cervical. Las ramas dorsales inervan los músculos y piel dorsal del cuerpo y las ventrales los músculos y partes ventrales del cuerpo incluyendo los miembros.
Cada nervio espinal posee fibras motoras o eferentes que transmiten impulsos del SNC a los músculos y órganos y fibras sensitivas aferentes que llevan impulsos al SNC. En consecuencia los nervios espinales poseen:
- Aferentes somáticos (estímulos externos)
- Aferentes viscerales (estímulos viscerales)
- Eferentes motores (llevan impulsos a los músculos esqueléticos)
- Eferentes viscerales (llevan impulsos a glándulas y músculos lisos)
Los nervios craneales parten del encéfalo y se distribuyen principalmente por cabeza y cuello. Poseen la misma estructura básica que los nervios espinales pero además pueden contener fibras eferentes viscerales que inervan musculaturas vinculadas a los aparatos respiratorio y digestivo, regulan la visión, la olfacción, gusto, funciones viscerales y motoras, etc.
Los pares craneales son los siguientes:
I.- Olfatorio
II.- Óptico
III.- Oculomotor o motor ocular común
IV.- Troclear
V.- Trigémino
VI.- Abducens o motor ocular externo
VII.- Facial
VIII.- Vestíbulo coclear o estatoacústico
X.- Vago
XI.- Accesorio
XII.- Hipogloso
Los ganglios nerviosos son masas grises que se componen de cuerpos neuronales y fibras nerviosas, son de diferentes tipos y podemos considerar:
Ganglios sensitivos: se hallan en las raíces dorsales de los nervios espinales y en la zona radicular de algunos nervios craneales.
Ganglios craneales.
Ganglios paravertebrales.
Ganglios prevertebrales.
Un ramo anastomótico es un manojo de fibras que pasa de un tronco nervioso a otro. Un plexo nervioso está formado por un intercambio de fibras de un nervio a otro adyacente. Se llama ramo comunicante a las ramas que ponen en conexión las ramas ventrales de los nervios espinales con ganglios o nervios espinales.
Sistema nervioso autónomo
La regulación y control de las funciones viscerales dependen del Sistema Nervioso Visceral, vegetativo o autónomo. Sus componentes se encargan de regular procesos vitales como la respiración, digestión, presión sanguínea, metabolismo, etc. Como estos procesos vitales se mantienen aún en los estados inconscientes tales como el sueño o bajo situación anestésica, esta parte del sistema nervioso tiene como principal característica su independencia funcional, de ahí la denominación de sistema nervioso autónomo. Consta de una porción llamada Sistema Simpático y otra llamada Sistema Parasimpático. Ambas están bajo control directo del hipotálamo que actúa como un centro integrador del SNC.
Estructura general: las fibras nerviosas autónomas poseen un cuerpo neuronal (llamado primera neurona) en el encéfalo o médula espinal y oto cuerpo neuronal (segunda neurona) en un ganglio fuera del SNC. De la primera neurona parten fibras hasta los ganglios autónomos (fibras preganglionares), allí hacen sinapsis con la segunda neurona y de ésta se desprenden las fibras postganglionares que llegan al órgano a inervar.
El sistema parasimpático
Representa la porción cráneo sacra del SNA. Consta de fibras preganglionares que emergen del encéfalo con ciertos nervios craneales y con los nervios espinales de la región sacra, estas fibras preganglionares hacen sinapsis en un ganglio autónomo y de allí parten las fibras postganglionares que van a los órganos efectores. Las fibras preganglionares parasimpáticas son largas y las posganglionares son cortas.
El sistema parasimpático caudal está compuesto por fibras preganglionares que emergen de la médula espinal a través del segundo, tercero y cuarto nervio sacro para formar después el nervio pélvico, el cual, retroperitonealmente, se dirige a los ganglios terminales de los plexos pélvicos (ganglio hipogástrico) así como a los plexos mioentérico y submucoso del colon descendente y del recto, teniendo aquí lugar la comunicación a la segunda neurona eferente. Las fibras posganglionares de los ganglios terminales inervan a la vejiga y glándulas genitales accesorias, facilitando la eliminación de su contenido, y, a través de los nervios erigentes, alcanzan los cuerpos cavernosos del pene y del clítoris donde determinan una acción vasodilatadora (erección).
El sistema simpático
Las neuronas simpáticas ocupan la zona más apical del asta lateral de la sustancia gris de la médula espinal en los segmentos torácicos y primeros lumbares, sistema simpático o toracolumbar, dan origen a fibras nerviosas eferentes, preganglionares simpáticas que salen a través de la raíz ventral del nervio espinal.
Las fibras simpáticas preganglionares (motoras) provenientes de la raíz ventral de los nervios espinales torácicos y lumbares llegan a los ganglios paravertebrales por intermedio del ramo comunicante blanco, aquí realizan la sinapsis o pueden continuar para realizarla en un ganglio prevertebral.
Las neuronas simpáticas más periféricas (externas) de esta columna establecen sinapsis en los ganglios paravertebrales de la cadena ganglionar simpática y ejercen su acción segmentariamente sobre arterias parietales, músculo erector del pelo y glándulas cutáneas (sudoríparas y sebáceas).
Su acción produce vasoconstricción periférica, erección pilosa e hiperhidrosis.
Las neuronas simpáticas situadas más profundas (internas) de esta columna establecen sinapsis en los ganglios prevertebrales y ejercen su acción sobre la capa muscular lisa de las arterias viscerales y los órganos que ésta irriga.
El efecto sobre estos órganos no es uniforme (acción inhibitoria o facilitadora).
Ganglios paravertebrales (pares)
Los ganglios paravertebrales se disponen en cadena a lo largo de la superficie lateroventral de los cuerpos vertebrales, se hallan unidos por fibras longitudinales para formar dos troncos simpáticos que se extienden desde la base del cráneo hasta los territorios coxígeos donde se reúnen en un solo ganglio.
La porción cervical contiene tres formaciones ganglionares conformadas por la fusión de los ocho ganglios segmentarios primitivos.
Ganglio cervical craneal: situado a la salida del cráneo.
Ganglio cervical medio: pequeño y puede estar ausente.
Ganglio cervical caudal: situado en la entrada del tórax, dorsolateralmente de la tráquea y medialmente de la 1° - 3° articulación costovertebral, se fusiona con los 3 o 4 primeros ganglios paravertebrales torácicos, formando el ganglio cérvicotoracico o ganglio estrellado.
La porción cervical del sistema simpático se completa con un tronco nervioso que partiendo del ganglio cervicotorácico, discurre unido al nervio vago formando el tronco vagosimpático, éste discurre dorsomedialmente a la arteria carótida común para terminar en el ganglio cervical craneal. De él se desprenden fibras simpáticas que pasan a formar parte de los nervios craneales.
Ganglios prevertebrales (impares)
Los ganglios prevertebrales (impares) son masas irregulares están situados en los plexos nerviosos mesentéricos que rodean a la aorta abdominal y sus principales ramas viscerales.
0 notes
Photo

3ra parte Mientras los ratones seguían saciados, los investigadores usaron técnicas genéticas para crear artificialmente el hambre al activar las neuronas que promueven el hambre en el hipotálamo. Estas neuronas expresan el gen para la proteína r-Agouti (AgRP) y se ha demostrado previamente que restaura comportamientos simples de la alimentación. Mediante la activación de estas neuronas AgRP, Livneh y sus colegas hicieron que los ratones saciados reaccionaran de nuevo a los estímulos visuales y buscaran más alimento y también restauraron el patrón de respuestas visuales de los alimentos en neuronas en la corteza insular como las observadas previamente en ratones hambrientos. "Estas neuronas AgRP causan hambre, son la quintaesencia de la neurona del hambre --explica Lowell--. Es un gran avance aprender que podemos activarlas artificialmente y hacer que ratones llenos trabajen para conseguir comida y comer como si no hubieran comido en mucho tiempo. Estas neuronas parecen capaces de provocar un conjunto diverso de comportamientos asociados con el hambre y con comer". Teniendo en cuenta su investigación, también puede ser posible marcar el camino específico de las neuronas AgRP a la corteza insular y reducir la sobre-atención a las señales de alimentos en el medio ambiente, sin afectar a la alimentación a la hora de las comidas. Sin embargo, los investigadores señalan que esta hipótesis requiere más investigación. Con su visión sin precedentes en la corteza insular, el equipo de Andermann y Lowell creó una hoja de ruta del circuito cerebral por el cual las neuronas AgRP relacionadas con el hambre finalmente influyen en la corteza insular. Usando poderosos métodos genéticos y ópticos para encender y apagar células individuales a voluntad, el equipo pudo observar los efectos tanto en las neuronas aguas abajo como en el comportamiento. El circuito que revelaron incluye la amígdala, que se cree que actualiza el valor de las señales de alimentos, y el tálamo paraventricular, que también es importante para los comportamientos motivados. Los autores sugieren que la vía puede sesgar la toma de decisiones aumentando los pros y disminuyendo las desventajas de buscar y co
0 notes
Text
Cómo se produce la percepción visual: del ojo al cerebro
La percepción visual es el resultado de múltiples interacciones complejas entre diferentes partes del SN. Las neuronas de cada estación de relevo del sistema visual
La retina envía la información al núcleo geniculado lateral dorsal del tálamo (NGLd) por medio del nervio óptico. Las fibras del nervio óptico se entrecruzan en el quiasmo óptico.
Las fibras que se originan en la parte nasal de cada retina cruzan en el hemisferio opuesto, mientras que las que se originan en la parte temporal proyectan directamente al hemisferio ipsilateral.
Esto significa que las fibras originadas en la retina temporal del ojo izquierdo y las originadas en la retina nasal del ojo derecho proyectan hacia el hemisferio izquierdo; y al revés, las fibras originadas en la retina nasal del ojo izquierdo y las originadas en la retina temporal del ojo derecho proyectan hacia el hemisferio derecho.
Óptica Alomar Barcelona's insight:
Mecanismo y proceso de la visión
La visión, fenómeno complejo que se recibe en el cortex cerebral (áreas visuales de la corteza occipital), donde se reconocen e interpretan las imágenes que llegan desde el ojo, ventana receptora de la información.
Los estímulos luminosos recogidos por el ojo van al cerebro donde se transforman en sensaciones visuales. El ojo ve y el cerebro interpreta el estímulo recibido.
0 notes
Text
Visión ciega: causas y síntomas del 'ver sin saber que se ve'
Sus ojos funcionan bien, están intactos. Pero dicen no ver nada. Y realmente ven, sin saber que ven. Este fenómeno tan curioso es lo que les ocurre a las personas que sufren de **visión ciega, un trastorno neurológico** provocado por un daño cerebral que afecta a la capacidad de representar conscientemente los estímulos visuales del entorno. En este artículo te explicamos qué es la visión ciega, cómo surje este concepto, cuáles son sus causas, y cómo diferenciarla de otros trastornos similares. * Artículo relacionado: "[Ceguera cortical: causas, síntomas y tratamiento](/clinica/ceguera-cortical)" ## Visión ciega: definición y antecedentes Visión ciega (*blindsight*) es un término acuñado por el psicólogo inglés, Lawrence Weiskrantz, que hace alusión a la capacidad que tienen algunos sujetos para detectar, localizar y discriminar estímulos visuales de forma inconsciente. **Las personas que padecen este trastorno “ven, sin saber que ven”**; es decir, no reconocen conscientemente los objetos que tienen delante, a pesar de que actúan como si, de facto, estuvieran ahí. Las primeras investigaciones sobre el fenómeno de la visión ciega se llevaron a cabo en animales, principalmente monos, con la extirpación quirúrgica de las regiones cerebrales encargadas de la visión (el área V1). Cuando se les privaba de estas estructuras, los animales parecían retener algunas capacidades visuales, como la habilidad para detectar el contraste o para diferenciar un objeto de otro en función de su forma. Pocos neurocientíficos creían que los seres humanos pudieran llegar a tener una visión normal con estas áreas cerebrales dañadas. Los pacientes cuya corteza corteza visual había sido destruida mostraban una ceguera total, o eso parecía. En 1973, el equipo del psicólogo alemán Ernst Pöppel comprobó que, aunque algunos de ellos carecían de corteza visual y declaraban que eran incapaces de ver los objetos, **los movimientos oculares de sus ojos se encaminaban hacia ellos**: era la evidencia de que su sistema visual estaba informando, de alguna manera, de la existencia de los mismos. Pero lo que terminó de convencer a la comunidad científica de que el fenómeno de la visión ciega merecía toda su atención fueron los trabajos de Larry Weiskrantz y sus colegas a comienzos de los años setenta. **En los experimentos se utilizó la técnica de elección forzada** (que obliga a los pacientes a escoger entre opciones definidas, en vez de preguntar únicamente qué es lo que ve): los pacientes tenían que elegir entre dos posibles colores o localizaciones, al tiempo que les pedían que adivinaran cuál era aplicable a un objeto visual que decían no poder ver. Las respuestas de algunos de los pacientes resultaron ser correctas en una proporción significativa; es decir, con una frecuencia mayor de lo que cabría esperar por azar. Fue a partir de entonces cuando se empezó a etiquetar a estas personas como pacientes con visión ciega. En la actualidad, se ha demostrado que las personas con visión ciega **no sólo pueden “intuir” el color o la localización de los objetos, sino también la orientación de líneas o enrejados, el momento de aparición o las expresiones de rostros**. Sin embargo, no pueden hacerlo con otros aspectos como la detección de matices sutiles o de movimientos complejos. ## Causas y estructuras cerebrales implicadas La visión ciega se presenta en una porción de nuestros órganos perceptivos: el escotoma o punto ciego. **Este fenómeno ocurre cuando hay un daño o una lesión en el lóbulo occipital, y más concretamente en la corteza visual primaria (V1)**, que se encarga del procesamiento de los estímulos visuales. Cuando recibimos información de un objeto a través de las retinas de nuestros ojos, ésta hace un recorrido que va desde las células ganglionares del nervio óptico hasta diversas estructuras subcorticales que, actuando a modo de zonas de relevo, se encargan de integrar la información de cada modalidad sensorial (en este caso, la vista). A nivel subcortical, la información visual transcurre por estructuras como el [bulbo raquídeo](/neurociencias/bulbo-raquideo), el mesencéfalo y el núcleo geniculado lateral del tálamo. **En este nivel, todavía no somos conscientes de lo que hemos “visto”**, ya que la información no ha llegado aún a los niveles corticales superiores. Sin embargo, ésta sí puede influir en nuestra conducta, tal y como ocurre en los casos de visión ciega, en los que la persona ve, sin saber que ve. Los pacientes con visión ciega tienen dañado, por tanto, el módulo final de un complejo circuito de procesamiento visual, que es insuficiente por sí mismo y sin el resto de estructuras sensoriales y subcorticales pero necesario, al mismo tiempo, para que exista un reconocimiento consciente de lo que percibimos. * Quizás te interese: "[Corteza visual del cerebro: estructura, partes y vías](/neurociencias/corteza-visual)" ## El modelo sensoriomotor de la visión El modelo convencional del fallo estructural en el procesamiento visual (que implica una lesión en varias áreas del cerebro) asume, de forma implícita, que la visión consiste en crear una representación interna de la realidad exterior, cuya activación generaría la experiencia visual consciente. No obstante, no es el único que se ha postulado para intentar explicar por qué se produce un fenómeno como el de la visión ciega. **El enfoque ecológico de la percepción visual que propuso el psicólogo James J. Gibson**, considera que la visión ha de entenderse como una herramienta necesaria para la supervivencia. Según Gibson, el valor real del procesamiento visual está en poder identificar y ver con la mirada qué hay y dónde, de modo que podamos evitar obstáculos, identificar alimentos o posibles amenazas, alcanzar metas, etc. Todo este trabajo de “deducción visual” lo realizaría la retina en interacción con múltiples señales ambientales. Y la clave estaría en **discriminar la información relevante, de entre tanta señal, para poder gestionar una conducta en particular**. En la actualidad, el enfoque de Gibson se ha reformulado como el modelo sensoriomotor de la visión, en el que se toman prestados conceptos del enfoque ecológico y se postula que la visión es una actividad para explorar nuestro entorno en base a las contingencias sensoriomotoras, no una representación que creamos internamente. ¿Qué quiere decir esto? Que **la visión no implica únicamente la recepción de información a través de nuestros ojos**; esa información se moldea y se transforma en función de los cambios motores (p. ej. los músculos de los ojos o la contracción pupilar) y sensoriales que acompañan dicha experiencia visual, así como por los atributos visuales de los propios objetos que percibimos. La diferencia básica entre el modelo sensoriomotor y el modelo convencional es que, este último, asume que si falla o falta una determinada región del cerebro (la corteza visual primaria) la representación interna desaparece de la percepción consciente, con lo que ello implica; al contrario, para el enfoque sensoriomotor, el mundo exterior no se recordaría en la mente de la persona que lo percibe y la realidad funcionaría como una memoria externa que se pone a prueba en las relaciones entre estímulos sensoriales y respuestas motoras. ## Diagnóstico diferencial A la hora de diagnosticarse, la visión ciega debe diferenciarse de otra serie de trastornos similares como la doble hemianopsia, la ceguera psíquica de Munk, la ceguera histérica y la ceguera simulada. ### Doble hemianopsia **El paciente tiene conservada la visión macular y central**, aunque tiene una visión en forma de “cañón de fusil”. Este trastorno puede preceder o seguir a la visión ciega. ### Ceguera psíquica de Munk La persona tiene dificultades para reconocer objetos (agnosia visual), aunque sí **conserva la sensación de conciencia visual**. ### Ceguera histérica **El paciente se muestra indiferente, pero sin anosognosia**. Los exámenes confirman que la visión es normal, a pesar de que la persona informa de sus problemas de visión parcial o total. ### Ceguera simulada **La persona se inventa su propia dolencia**, en este caso la ceguera, para asumir el papel de enfermo (síndrome de Münchhausen) #### Referencias bibliográficas: * Aldrich MS, Alessi AG, Beck RW, Gilman S. Cortical blindness: etiology, diagnosis and prognosis. Ann Neurol 1987; 21: 149 - 158. * Brogaard, B. (2011). Are there unconscious perceptual processes. Consciousness and Cognition, 20, 449-463. * O'Reagan, J. & Noë, A. (2001). A sensorimotor account of vision and visual consciousness. Behavioral and Brian Sciences, 24, 939 - 973.
Ver Fuente
Ver Fuente
0 notes
Text
Reflejo fotomotor: qué es y cómo funciona esta reacción de la pupila
El reflejo fotomotor es un automatismo de nuestro sistema nervioso que nos protege de los cambios en la intensidad y del exceso de luz. Su función es hacer reaccionar a la pupila para que reduzca o aumente su tamaño, de modo que permita que llegue a nuestros ojos la cantidad adecuada de luz del ambiente. **En este artículo te explicamos qué es el reflejo oculomotor** y cómo actúa, de qué está compuesto el circuito responsable de este reflejo, cuáles son las principales funciones que lleva a cabo y de qué modo se evalúa clínicamente. * Artículo relacionado: "[Sistema nervioso autónomo: estructuras y funciones](/neurociencias/sistema-nervioso-autonomo)" ## ¿Qué es el reflejo fotomotor? El reflejo fotomotor se produce **cuando la pupila reacciona y se contrae o se dilata como respuesta a un estímulo lumínico**. Este arco reflejo gestionado por el sistema nervioso autónomo nos sirve para controlar que la cantidad de luz a la que se exponen nuestros ojos es la adecuada, de modo que se evite una sobreexposición o un deslumbramiento. En personas sanas, el aumento del diámetro de la pupila se conoce con el nombre de midriasis y es una reacción normal que se produce cuando hay poca luz o penumbra; al contrario, la contracción pupilar se denomina miosis y ocurre cuando hay un aumento de la luminosidad. El reflejo fotomotor y el consecuente cambio de tamaño en las pupilas es bilateral y se produce de forma simultánea en ambos ojos cuando alguno de ellos recibe el estímulo lumínico; no obstante, **recibe el nombre de reflejo fotomotor directo cuando se contrae la pupila en el ojo que recibe el estímulo; y reflejo fotomotor consensual cuando la pupila que se contrae es la del ojo contrario**. La tarea de controlar las variaciones en el tamaño pupilar la llevan a cabo dos músculos oculares: el esfínter de la pupila, que se encarga de la contracción a través de las denominadas fibras parasimpáticas; y el músculo dilatador, situado en la zona posterior del iris, es el responsable de dilatar las pupilas y está controlado por fibras del sistema nervioso simpático. * Quizás te interese: "[Las 11 partes del ojo y sus funciones](/neurociencias/partes-del-ojo)" ## Estructura y fisiología El correcto funcionamiento del reflejo fotomotor depende de todas y cada una de las partes implicadas en el circuito de dicho arco reflejo. Veamos, a continuación, cuáles son: ### 1. Fotorreceptores Los receptores encargados de iniciar el reflejo fotomotor **pertenecen a las células de la retina especializadas en la percepción de los estímulos lumínicos.** Los fotorreceptores clásicos son los conos, responsables de la percepción del color; los bastones o bastoncillos, encargados de la visión en condiciones de baja visibilidad; y las células ganglionares de la retina, cuya función es transmitir los impulsos que inician el arco fotomotor a través de neuronas intermediarias. Cuando la luz estimula las células fotorreceptoras se produce un proceso de transducción que convierte los estímulos lumínicos en impulsos eléctricos que se transmiten hacia las zonas del cerebro encargadas de procesar la visión a través de vías aferentes. ### 2. Vías aferentes Una vez que el estímulo lumínico ha incidido sobre la retina viajará a través de una vía aferente, las fibras sensitivas del nervio oftálmico, hasta el sistema nervioso central; y desde ahí, una parte de las fibras nerviosas especializadas del nervio óptico se separan y transmiten la información al mesencéfalo. El resto de fibras transmiten la información y hacen relevo en los cuerpos geniculados, situados en la cara posterior del tálamo, para después dirigirse a la corteza visual primaria. Con todo, cabe señalar que **el reflejo motor se integra en el mesencéfalo sin que se produzca una intervención de niveles funcionales superiores**, lo que indica que en los casos en los que hay un daño a nivel de los cuerpos geniculados o la corteza visual, este arco reflejo no se vería afectado. * Quizás te interese: "[Reflejo de Moro: características e implicaciones clínicas en los bebés](/desarrollo/reflejo-de-moro)" ### 3. Núcleos de integración Toda vez que las fibras nerviosas sensitivas que provienen del nervio óptico alcanzan el mesencéfalo, **llegan al pretectum o área pretectal del mismo, que se localiza justo delante de los colículos superiores y por detrás del tálamo**. Las fibras que provienen del nervio óptico transmiten la información a dos núcleos ganglionares: el núcleo del tracto visual y el núcleo olivar. La información acerca de la intensidad lumínica se procesa en estos núcleos. Después, a través de interneuronas se conectan el núcleo olivar y el tracto visual con el núcleo de Edinger-Westphal, desde donde salen las fibras motoras simpáticas que inducen el movimiento y la respuesta efectora. ### 4. Vías eferentes Los axones del sistema nervioso simpático emergen desde el núcleo de Edinger-Westphal hacia la órbita, junto con las fibras del nervio fotomotor. Una vez que este último llega a la órbita, **las fibras simpáticas salen y alcanzan el ganglio ciliar**, que actúa como última estación de relevo en la integración del reflejo fotomotor, y desde donde emergen los nervios ciliares cortos que se encargan de la inervación simpática del ojo. ### 5. Efectores Finalmente, los nervios ciliares cortos inervan el músculo ciliar, y mediante su estimulación provocan que se contraiga y, consecuentemente, **se produce la contracción pupilar**. Así, el músculo ciliar se encarga de que la pupila reduzca su tamaño y permita que entre menos luz en el ojo. ## Funciones Una de las principales funciones del reflejo fotomotor es **asegurar que la cantidad de luz que ingresa en el ojo es la adecuada**: ni demasiada luz, lo que provocaría un deslumbramiento; ni luz insuficiente, ya que las células fotorreceptoras no podrían ser estimuladas correctamente y la visión sería deficiente. Cuando se produce un exceso en la absorción de estímulos lumínicos, la transducción que se genera en las células fotorreceptoras es inadecuada, las reacciones químicas ocurren demasiado rápido y los precursores se consumen antes de que puedan regenerarse, derivando en un deslumbramiento o una sobreexposición a la luz. El efecto de deslumbramiento es el que se produce, por ejemplo, cuando pasamos de un ambiente muy oscuro o de tener los ojos cerrados a abrirlos y encontrarnos con una fuente de luz muy intensa. **Lo que ocurre es que nos ciega y somos incapaces de ver durante unos cuantos segundos**, hasta que las células de la retina se acomodan a la intensidad de luz ambiental. Aunque la función del reflejo fotomotor es, precisamente, evitar que se produzca esa sobreexposición a la luz, lo cierto es que en ocasiones no es suficiente y el efecto se produce igualmente debido a que hace falta cierto tiempo para que el estímulo lumínico se convierta en impulso eléctrico y se produzca el arco reflejo, y la posterior contracción pupilar. ## Evaluación clínica del reflejo **La evaluación clínica del reflejo fotomotor suele realizarse con la ayuda de una linterna**. Se proyecta la luz al ojo con el fin de ver cómo reacciona la pupila y, en el caso de que ésta disminuya de tamaño en respuesta al estímulo lumínico, tendremos una pupila normorreactiva; si, por el contrario, la pupila reacciona débilmente ante la luz, tendremos una pupila hiporreactiva. Otro de los objetivos de la evaluación de este arco reflejo es conocer si existe algún tipo de daño o lesión en el nervio óptico, así como comprobar si hay pérdida de la visión. Durante la exploración, también suele ser habitual comprobar si el reflejo consensual está intacto: esto se hace observando si se contrae la pupila del ojo contrario al que está siendo estimulado por la luz. Por último, si durante la exploración se observa cualquier reacción anormal de la pupila a la estimulación lumínica, **es importante evaluar otros aspectos del sistema visual por si existieran daños en otras vías nerviosas** del sistema visual, más allá del reflejo fotomotor. #### Referencias bibliográficas: * Hultborn, H., Mori, K., & Tsukahara, N. (1978). The neuronal pathway subserving the pupillary light reflex. Brain Research, 159(2), 255 - 267. * Kaufman, P. L., & Alm, A. (Eds.). (2004). Adler fisiología del ojo: aplicación clínica. Elsevier. * McDougal, D. H., & Gamlin, P. D. (2010). The influence of intrinsically-photosensitive retinal ganglion cells on the spectral sensitivity and response dynamics of the human pupillary light reflex. Vision research, 50(1), 72 - 87.
Ver Fuente
Ver Fuente
0 notes
Text
Neuroetología: ¿qué es y qué investiga?
La neuroetología es una ciencia que se encarga de estudiar el comportamiento animal desde el punto de vista del sistema nervioso. Para ello, se sirve de dos disciplinas científicas: la neurobiología, que estudia la organización y la estructura de las células nerviosas y su relación con la conducta; y la etología, la rama de la ciencia que estudia el comportamiento de los animales en su hábitat natural. En este artículo te explicamos **qué es la neuroetología, cuáles son sus orígenes, así como sus científicos más relevantes** y las principales contribuciones a esta disciplina. * Artículo relacionado: "[¿Qué es la Etología y cuál es su objeto de estudio?](/psicologia/etologia)" ## ¿Qué es y qué estudia la neuroetología? La neuroetología es una **disciplina científica que estudia, desde un enfoque evolutivo y comparado, los mecanismos neurofisiológicos implicados en el comportamiento animal**. Esta neurociencia relativamente nueva se centra en aplicar estrategias que permiten desentrañar qué estructuras y funciones del sistema nervioso están implicadas en el despliegue de las conductas típicas de las distintas especies animales. Como su propio nombre indica, esta disciplina aúna los conocimientos de la neurobiología, que estudia el funcionamiento y la organización de las células del sistema nervioso que median en el comportamiento; y **la etología, la rama de la ciencia que se ocupa de indagar en cómo y por qué los animales se actúan como lo hacen**. El neurocientífico alemán Jörg-Peter Ewert, uno de los pioneros en la investigación neuroetológica, considera que esta ciencia tiene como objetivo intentar responder a preguntas relevantes, como qué mecanismos fisiológicos explican la conducta; de qué forma se desarrolla dicha conducta en el animal; cómo una determinada conducta favorece la adaptación de un ejemplar y su descendencia, o cuál es la historia filogenética de una determinada conducta. Los especialistas en neuroetología **utilizan en sus estudios científicos animales con habilidades singulares que puedan servir de modelo comparativo** a la hora de estudiar las propiedades de su sistema nervioso y cómo éste ha sido capaz de adaptarse a lo largo del desarrollo ontogenético (y a nivel filogenético, en toda su especie) para poder desplegar determinados comportamientos que se ajustan a contextos específicos. ## Antecedentes: la etología y las 4 preguntas de Tinbergen Como hemos comentado anteriormente, la neuroetología se nutre de la etología, que se encarga de estudiar el comportamiento animal. Una de las mayores aportaciones a esta disciplina la hizo durante la primera mitad del siglo pasado el zoólogo holandés, Nikolaas Tinbergen, considerado como uno de sus máximos exponentes. **Tinbergen entendía el comportamiento de los animales como patrones motores estereotipados** que están controlados tanto por mecanismos fisiológicos internos, como por determinados estímulos externos. Según él, cada animal estaría dotado de una maquinaria de comportamiento estrictamente limitada y altamente compleja, la cual es constante a través de una especie o población. La clave para Tinbergen, y para los etólogos en general, era responder a la pregunta de por qué los animales se comportan como lo hacen y, a su vez, intentar comprender las siguientes claves relacionadas con dicha cuestión: ### ¿Cuál es el mecanismo de control que subyace a la conducta animal? La respuesta a esta pregunta implicaría **tener en cuenta factores internos (hormonales o neurales) y externos** (estímulos táctiles o visuales, p. ej.). ### ¿Cómo se logra dicha conducta en el desarrollo ontogenético del ejemplar animal? Esta respuesta supondría indagar en la historia del animal, **determinando las posibles influencias genéticas y ambientales que han favorecido dicho desarrollo**. ### ¿Cuál es el valor adaptativo o de supervivencia de determinada conducta? En esta respuesta habría que tomar en consideración aspectos como el significado evolutivo o la ventaja selectiva de determinados comportamientos animales. ### ¿Cómo ha evolucionado dicha conducta en el curso de la historia de la especie animal? En este caso, la respuesta supondría **dar cuenta de la historia evolutiva de la conducta en cuestión**, analizando los factores evolutivos necesarios para la aparición del comportamiento en la especie. * Quizás te interese: "[Neurociencias: la nueva forma de entender a la mente humana](/neurociencias/neurociencias-mente-humana)" ## Aportaciones de la neurobiología **La neurobiología, que estudia los mecanismos biológicos mediante los cuales el sistema nervioso regula el comportamiento**, es otra de las disciplinas científicas de las que se nutre la neuroetología. Esta ciencia debe su origen, principalmente, a una serie de avances técnicos y teóricos, en materia de investigación del sistema nervioso, que se produjeron también a mediados del siglo XX. En este sentido, fueron varios los hitos que marcaron el auge de la neurobiología: la aparición de la doctrina de la neurona de Ramón y Cajal, la presentación del modelo de potencial de acción de Hodgkin y Huxley, así como el desarrollo de técnicas histológicas, de estimulación, registro y trazado de conexiones neurales. **Estos avances facilitaron que en la década de los 70 Tinbergen hiciera un llamamiento en favor de una síntesis entre la etología y la neurobiología** para dar paso a la neuroetología, aunque al principio no fue fácil dado que hubo grandes dificultades para conseguir los métodos adecuados para relacionar las actividades de neuronas o tejidos neurales individuales con la conducta del animal en su hábitat natural. Finalmente, **fueron varios los científicos que impulsaron el desarrollo de la neuroetología**; por ejemplo, Erich von Holst, con su técnica de estimulación cerebral focal, consiguió demostrar mediante varios experimentos con animales que la estimulación de ciertas áreas del cerebro de los pollos podía provocar patrones de conducta agresiva; o Karl Von Frisch, Hansjochem Autrum y Ted Bullock, que fueron pioneros en la investigación sobre los aspectos neurofisiológicos que subyacen al comportamiento específico de abejas, artrópodos y peces. ## Las investigaciones de Jorg-Peter-Ewert Como hemos comentado al inicio, Jorg-Peter-Ewert es uno de los científicos que más ha investigado en el campo de la neuroetología. Uno de sus estudios más relevantes se llevó a cabo con anfibios, concretamente con ejemplares de sapo común, con el objetivo de **investigar los mecanismos neurales implicados en la capacidad de respuesta selectiva de estos animales**. El científico alemán estudió dos tipos de comportamientos controlados visualmente: los de orientación o captura de presas y los de evitación o escape. Para elicitar respuestas de captura utilizó estímulos visuales como barras largas, delgadas y oscuras (simulando ser lombrices), que provocaban una rápida reacción en el sapo al moverse a través de sus retinas. En el momento en el que se añadían dos barras laterales al estímulo (en forma de “ele”), la respuesta de ataque del sapo tendía a inhibirse (la “lombriz” pasó a ser una “no lombriz”). **Las células ganglionares de la retina del animal respondían a objetos con la velocidad y la forma adecuadas**, pero eran incapaces de discriminar entre el estímulo lombriz y no lombriz. En el estudio se halló que estimulando una estructura denominada tectum óptico se generaba el patrón conductual asociado a la respuesta de ataque del sapo; y por otra parte, mediante la estimulación eléctrica de determinadas partes del tálamo se provocan respuestas defensivas de escape y huida. Esta investigación es solo un ejemplo de lo que en su día fue un estudio pionero que aportó grandes conocimientos a la neuroetología. En la actualidad, se sigue investigando la conducta animal y sus correlatos neuronales en numerosas especies animales: desde la conducta de agresión en aves, el comportamiento depredador de las lagartijas o la conducta social de los primates. #### Referencias bibliográficas: * Ewert, J. P. (1987). Neuroethology of releasing mechanisms: prey-catching in toads. Behavioral and Brain Sciences, 10(3), 337-368. * Pflüger, H. J., & Menzel, R. (1999). Neuroethology, its roots and future. Journal of Comparative Physiology A, 185(4), 389-392.
Ver Fuente
Ver Fuente
0 notes
Last Seen Blogs

jihyesx-blog
이지혜

ariesdevilxoxo
moonlight

hmprgrbrg
Adj egy cigit, eskuszom buszra kell.

fullyarmedhetalian
multi-fandom mess

ekarmaindia2001
EKARMAINDIA