
leaping-laelaps-art
A Konservat-Lagerstätte enthusiast's art blog
The art-bloggening
23 posts
Don't wanna be here? Send us removal request.
Last Seen Blogs

hidinhaydn
trying my damndest

phsussa
Sem título

faejilly
half agony, half hope

shattered-burning-soul
~Shattered Soul~

igowiththemoon-sayinggoodnight
the moon made me do it
Text
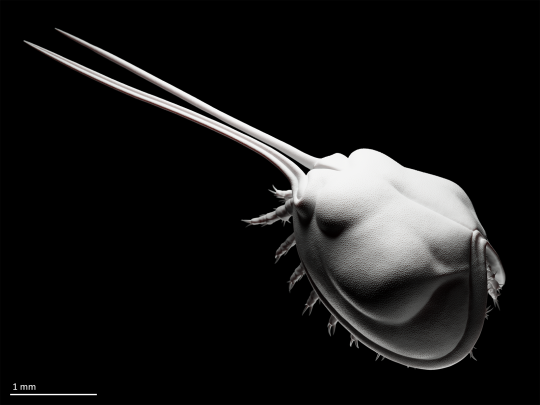
Sculpt of the small bivalved arthropod Gladioscutum lauriei from the middle Cambrian of Australia (after Hinz-Schallreuter & Jones 1994).
Gladioscutum had a body only 2-3 mm long, but, being undoubtedly aware of its disappointingly small size compared to its cooler Cambrian cousins like radiodonts and trilobites, tried to make up for it with a pair of (presumably) front-facing spines that were at least as long as the rest of the head shield.
Other than improving its self-esteem, the function of Gladioscutum's extremely elongated spines is unknown. The enlarged spines of other small Cambrian bivalved arthropods have been suggested to fulfill a sensory role, but this remains speculative (Zhang et al. 2014).
References and notes:
Gladioscutum was originally described as an "archaeocopid", an order that is now known to be an artificial grouping of various small bivalved arthropod fossils superficially resembling modern ostracod crustaceans. To my knowledge, the affinities of Gladioscutum have not been reinvestigated since its initial description, but its appearance (marginal rims, valve lobation, ornamented surface, simple hinge line) and age seem bradoriid-y enough (Hou et al. 2001) for me to more or less confidently reconstruct it as one (top scientific rigour as always on this blog).
Appendage morphology is unknown in Gladioscutum - what little soft anatomy I have not modestly hidden under the head hield is based on the bradoriid Indiana sp. from the Chengjiang Biota (Zhai et al. 2019). In that species, only the antennae are differentiated from the rest of the appendages, which has the double advantage of (1) not making crazy hypotheses about limb specialization in Gladioscutum and (2) giving me fewer different types of limbs to sculpt.
Like Gladioscutum, most bradoriids are only known from their decay-resistant valves, which are often squashed flat in a so-called "butterfly" position. This arrangement has been traditionally interpreted as the life position of the animals, which were implied to crawl over the seafloor like tiny crabs (e.g., Hou et al. 1996). Yet, undistorted fossils of head shields preserved in 3D are almost always closely drawn together, which is similar to the way modern bivalved arthropods like ostracods are articulated (protecting the soft limbs and body) and probably more reflective of the actual life position of bradoriids (Betts et al. 2016), as depicted here.
References:
Betts, M. J., Brock, G. A., & Paterson, J. R. (2016). Butterflies of the Cambrian benthos? Shield position in bradoriid arthropods. Lethaia, 49(4), 478–491. https://doi.org/10.1111/let.12160
Hinz-Schallreuter, I., & Jones, P. J. (1994). Gladioscutum lauriei n.gen. N.sp. (Archaeocopida) from the Middle Cambrian of the Georgina Basin, central Australia. Paläontologische Zeitschrift, 68(3), 361–375. https://doi.org/10.1007/BF02991349
Hou, X., Siveter, D. J., Williams, M., Walossek, D., & Bergström, J. (1997). Appendages of the arthropod Kunmingella from the early Cambrian of China: Its bearing on the systematic position of the Bradoriida and the fossil record of the Ostracoda. Philosophical Transactions of the Royal Society of London. Series B: Biological Sciences, 351(1344), 1131–1145. https://doi.org/10.1098/rstb.1996.0098
Hou, X., Siveter, D. J., Williams, M., & Xiang-hong, F. (2001). A monograph of the Bradoriid arthropods from the Lower Cambrian of SW China. Earth and Environmental Science Transactions of The Royal Society of Edinburgh, 92(3), 347–409. https://doi.org/10.1017/S0263593300000286
Zhai, D., Williams, M., Siveter, D. J., Harvey, T. H. P., Sansom, R. S., Gabbott, S. E., Siveter, D. J., Ma, X., Zhou, R., Liu, Y., & Hou, X. (2019). Variation in appendages in early Cambrian bradoriids reveals a wide range of body plans in stem-euarthropods. Communications Biology, 2(1), Article 1. https://doi.org/10.1038/s42003-019-0573-5
Zhang, H., Dong, X., & Xiao, S. (2014). New Bivalved Arthropods from the Cambrian (Series 3, Drumian Stage) of Western Hunan, South China. Acta Geologica Sinica - English Edition, 88(5), 1388–1396. https://doi.org/10.1111/1755-6724.12306
#i'll probably make more art of cambrian bivalved arthropods with fucked up spines#because there is no shortage of them#and they still manage to all look rather distinct#gladioscutum#bradoriid#(?)#arthropod#cambrian#paleozoic#australia#paleontology#palaeoblr#paleoart#my art
34 notes
·
View notes
Text

Falcatamacaris bellua is not, as it may seem at first glance, a fictive trilobite designed by a 6-year old, but a large (~10 cm) arthropod from the Weeks Formation (early Cambrian) notably distinguished by curved pleural spines (giving it its name*), a weakly biomineralized cuticle, and an unwillingness to be classified precisely.
Bonus views of the quick 3D model I made as a drawing reference:


References and notes:
*name which, by the way, is based on an incorrect use of the latin falcatus,-a,-um in its unexplainably inflected form falcatam, which exasperates me disproportionately (name should be Falcatacaris (how did the reviewers not give them shit for that mistake?)) (i am experiencing several taxonomy-related annoyances these days (perhaps i just need something inconsequential on which to take out my anger (anyway))).
Available fossil material of Falcatamacaris only features the dorsal carapace along with limited evidence for 3 cephalic pairs of limbs (but apparently no antennae) (Ortega-Hernández et al. 2015). The walking appendage morphology and arrangement depicted here is therefore only speculative and based on a generalized artiopodan (a broad group including trilobites and friends, within which Falcatamacaris has an uncertain position), after Sein & Selden (2012).
References:
Ortega-Hernández, J., Lerosey-Aubril, R., Kier, C., & Bonino, E. (2015). A rare non-trilobite artiopodan from the Guzhangian (Cambrian Series 3) Weeks Formation Konservat-Lagerstätte in Utah, USA. Palaeontology, 58(2), 265–276. https://doi.org/10.1111/pala.12136
Stein, M., & Selden, P. A. (2012). A restudy of the Burgess Shale (Cambrian) arthropod Emeraldella brocki and reassessment of its affinities. Journal of Systematic Palaeontology, 10(2), 361–383. https://doi.org/10.1080/14772019.2011.566634
#mild taxonomic pissiness#just another Junnn11-inspired piece before getting back to serious business#as a treat#falcatamacaris#artiopod#arthropod#cambrian#paleozoic#paleontology#palaeoblr#paleoart#my art
48 notes
·
View notes
Text
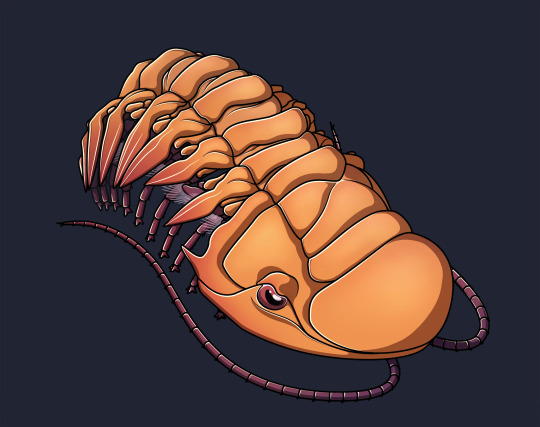
The Devonian trilobite Crotalocephalina gibba.
Made as a more or less successful exercise to emulate the style of Junnn11, the person who illustrated many of the wikipedia articles on Cambrian arthropods and Paleozoic euchelicerates (and whose art I happen to like very much).
#very accidental lesbian color scheme#i swear i didn't do it on purpose the colors just looked nice#i only realized two hours after picking the colors and went with it#it's not my fault if lesbians have the best pride flag#(perhaps with the exception of the aro and aroace ones - but there certainly is no reason for my opinion on these to biased hehe)#anyway#drawing based on a fossil i have on my desk#minus the appendages of course (those are mainly adapted from Ceraurus pleurexanthemus)#because there's no way on earth i'm drawing a cheirurid in a non-neutral pose with just my imagination and a few top view photos/diagrams#all these furrows and little knobs around the articulations!!!#also heeeeeyy i'm not dead i'm still making art#between work travel and the fact that i've started sketching on paper again over the last 6 months#i haven't done much digital art worth posting#(still hesitating to post my paper sketches on here)#but i do have some cool 3d projects in the works#so i will probably be posting some pieces in my usual style in the more or less near future#crotalocephalina#cheirurid#trilobite#arthropod#devonian#paleozoic#paleontology#palaeoblr#paleoart#my art
15 notes
·
View notes
Text


Europasaurus holgeri, a macronarian sauropod from the Late Jurassic of Germany, most famous for its unusually small size (just over 6 m long for mature specimens) attributed to insular dwarfism.
(Repost of some older 2022 art from my general-purpose blog, with additional renders and a proper bibliography)
References:
Marpmann, J. S., Carballido, J. L., Sander, P. M., & Knötschke, N. (2015). Cranial anatomy of the Late Jurassic dwarf sauropod Europasaurus holgeri (Dinosauria, Camarasauromorpha): Ontogenetic changes and size dimorphism. Journal of Systematic Palaeontology, 13(3), 221–263. https://doi.org/10.1080/14772019.2013.875074
Martin Sander, P., Mateus, O., Laven, T., & Knötschke, N. (2006). Bone histology indicates insular dwarfism in a new Late Jurassic sauropod dinosaur. Nature, 441(7094), Article 7094. https://doi.org/10.1038/nature04633
#europasaurus#macronaria#sauropod#dinosaur#reptile#vertebrate#jurassic#mesozoic#paleontology#palaeoblr#paleoart#my art
35 notes
·
View notes
Text


The archeocete Perucetus colossus dives through a coastal bloom of jellyfish in the Pisco Basin (southern Peru), some time during the Eocene (with bonus multiview).
I originally intended to add epibionts to this reconstruction (reflecting the specialized communities found on many living whales, especially baleen whales). Yet, interestingly, it appears that most animal epibionts and ectoparasites of modern cetaceans, such as whale barnacles (Hayashi et al. 2013) and remoras (Friedman et al. 2013), only appeared in the Neogene or late Paleogene, or have a poorly known (co-)evolutionary history, like whale lice (Pfeiffer 2009, Iwasa-Arai & Serejo 2018) and pennellids (large parasitic copepods) (Hermosilla et al. 2015). So, no epibionts* for big lad Perucetus!
References and notes about the reconstruction:
*animal epibionts. Unicellular eukaryotes like diatoms were most likely present on early cetaceans, given their prevalence on modern large marine animals (Ashworth et al. 2022). Of course, it is possible that other animals (i.e., early, less specialized representatives of modern groups, or different taxa altogether) were also already exploiting the surfaces offered by these early whales; however, this remains entirely speculative.
The reconstruction of Perucetus proposed in its original description (Bianucci et al. 2023) includes some rather odd (if interesting) choices about soft tissues, including limbs with webbed and distinguishable fingers, and a manatee-like tail. While these choices might be defendable in light of the rather basal status of Perucetus among cetaceans, I opted for a more derived look based on the assumption that fully marine cetaceans like basilosaurids would have probably rapidly acquired hydrodynamically favorable adaptations, pushing them towards a more familiar Neoceti-like appearance (even though Perucetus itself was likely a poor swimmer (Bianucci et al. 2023), it seems likely to me that this was a secondarily acquired trait, given the less extreme morphology of other basilosaurids).
Reconstruction in the multiview scaled to ~18 m in length after the estimations of Bianucci et al. (2023).
References:
Ashworth, M. P., Majewska, R., Frankovich, T. A., Sullivan, M., Bosak, S., Filek, K., Van de Vijver, B., Arendt, M., Schwenter, J., Nel, R., Robinson, N. J., Gary, M. P., Theriot, E. C., Stacy, N. I., Lam, D. W., Perrault, J. R., Manire, C. A., & Manning, S. R. (2022). Cultivating epizoic diatoms provides insights into the evolution and ecology of both epibionts and hosts. Scientific Reports, 12(1), Article 1. https://doi.org/10.1038/s41598-022-19064-0
Bianucci, G., Lambert, O., Urbina, M., Merella, M., Collareta, A., Bennion, R., Salas-Gismondi, R., Benites-Palomino, A., Post, K., de Muizon, C., Bosio, G., Di Celma, C., Malinverno, E., Pierantoni, P. P., Villa, I. M., & Amson, E. (2023). A heavyweight early whale pushes the boundaries of vertebrate morphology. Nature, 620(7975), Article 7975. https://doi.org/10.1038/s41586-023-06381-1
Friedman, M., Johanson, Z., Harrington, R. C., Near, T. J., & Graham, M. R. (2013). An early fossil remora (Echeneoidea) reveals the evolutionary assembly of the adhesion disc. Proceedings of the Royal Society B: Biological Sciences, 280(1766), 20131200. https://doi.org/10.1098/rspb.2013.1200
Hayashi, R., Chan, B. K. K., Simon-Blecher, N., Watanabe, H., Guy-Haim, T., Yonezawa, T., Levy, Y., Shuto, T., & Achituv, Y. (2013). Phylogenetic position and evolutionary history of the turtle and whale barnacles (Cirripedia: Balanomorpha: Coronuloidea). Molecular Phylogenetics and Evolution, 67(1), 9–14. https://doi.org/10.1016/j.ympev.2012.12.018
Hermosilla, C., Silva, L. M. R., Prieto, R., Kleinertz, S., Taubert, A., & Silva, M. A. (2015). Endo- and ectoparasites of large whales (Cetartiodactyla: Balaenopteridae, Physeteridae): Overcoming difficulties in obtaining appropriate samples by non- and minimally-invasive methods. International Journal for Parasitology: Parasites and Wildlife, 4(3), 414–420. https://doi.org/10.1016/j.ijppaw.2015.11.002
Pfeiffer, C. J. (2009). Whale Lice. In W. F. Perrin, B. Würsig, & J. G. M. Thewissen (Eds.), Encyclopedia of Marine Mammals (Second Edition) (pp. 1220–1223). Academic Press. https://doi.org/10.1016/B978-0-12-373553-9.00279-0
#'a heavyweight early whale pushes the boundaries of...' blablabla you've all read it by now#i have nothing to add#it's fat#look at it#that is all#perucetus#cetacean#mammal#vertebrate#eocene#cenozoic#paleontology#palaeoblr#paleoart#my art
399 notes
·
View notes
Note
Just wanted to say that your art, especially the 3D models, are just absolutely, unironically great to me. I love how there is obviously so much care and knowledge put into it and now really need to see whether I can get access to those papers so I can read more about the cool sponges and the Emu Bay Shale and just the lime stone pebble, even if I don't understand all of it.
It might sound weird because I can't describe why, but your art is really exciting in the "science can be so much fun" way. You obviously know what you're doing in both regards and get together the Looks amazing and Communicates how interesting it is. I know I discovered this blog like yesterday but I can't stop coming back to it.
Looking forward to whatever you're going to do in the future and wish you all the best!
Thank you so much for your kind words! There really is no greater compliment to me than being told that my art conveys my love and excitement for paleontology in a way that is both informative and visually pleasing!
My illustrations are usually strongly influenced by my research work (since I'm apparently part of this masochistic breed that does want to work in academia). In fact, most of my inspiration comes when reading a paper and thinking "isn't that neat?". Paleoart is such an efficient medium for communicating paleontological ideas to all audiences (experts and general public alike), and that's the way I approach all my pieces - a kind of graphical abstract, if you will. I'm glad the message comes across!
In addition to this mostly didactic concept of paleoart, I also love making it because the reconstruction process forces to ask certain questions in order to fill visual gaps (from mundane things like coloration, to more significant elements like missing anatomy or life environment) and pushes to study many different aspects of the fossils (e.g., phylogenetic affinities, functional morphology, taphonomy, sedimentology, etc.). I always learn a lot from this exploration, and some of the questions may not have ever been seriously asked before, which can lead to some pretty interesting (and fun) reflections (does anyone have any idea what colors would be most likely for an archaeocyath?).
In any case, I'm so happy you seem to be as excited about looking at my art as I am about making it! More is certainly coming, and now that I have a bit of time I hope to be able to make some more often.
A last note regarding papers: if you (or anyone) would like to read a paper I cited but cannot access it, just send me a DM and I'll be happy to share the pdf with you if I can!
#ask#“science can be so much fun” is very much the feeling i seek to instill into all my pieces#thank you for getting this and putting it into words#saving this ask to some place where i can look at it again from time to time#because it's the sweetest thing anyone has ever told to me about my art and really understands where i'm trying to go with it
2 notes
·
View notes
Text

I do not usually post WIPs, but I am happy with this one. The newly described basilosaurid Perucetus colossus (i.e., Chonkocetus the Large), contender for the title of heaviest known animal.
Missing parts (i.e., most of the animal, as customary for fossil vertebrates) based on Basilosaurus spp., Dorudon atrox, modern whales (Neoceti), and sirenians.
#will include proper references in the post for the finished piece#perucetus colossus#perucetus#cetacean#mammal#vertebrate#eocene#cenozoic#paleontology#palaeoblr#paleoart#my art#wip
279 notes
·
View notes
Text
I'm happy to report that this wonderful radiodont fellow finally has a proper name: Echidnacaris briggsi!
Published yesterday - along with the other radiodont from the Emu Bay Shale, now known as Anomalocaris daleyae - in this paper (which is unfortunately not open access, but I'm sure it will get shared somewhere...)

514 millions years ago in what will one day be known as the Emu Bay Shale (South Australia), a tiny Isoxys glaessneri encounters the hunter 'Anomalocaris' briggsi.
'Anomalocaris' briggsi was a large suspension-feeding radiodont related to the famous raptorial predator Anomalocaris canadensis. It is one of two radiodont species for which exceptionally detailed fossils of compound eyes are known. The eyes in this species in this species are unsual for radiodonts in that they are not stalked, and protected by a small plate which was likely a modified version of the lateral carapace elements found in hurdiids. The eye morphology suggests that 'A.' briggsi was a mesopelagic species capable of inhabiting depths of several hundred meters, using its acute vision to detect planktonic prey (Paterson et al. 2020).
Isoxys was a cosmopolitan genus of stem-euarthropod in the Lower and Middle Cambrian, characterized by a bivalved shield covering its whole body, two large eyes, and a frontal pair of so-called 'great appendages' probably used for grasping food items. These appendages show similarity with both the frontal appendages of megacheirans and those of radiodonts like Anomalocaris, and its mix of derived and basal anatomical traits (such as biramous appendages but an unclerotized trunk) make it a crucial organism for understanding the early evolution of arthropods (Legg & Vannier 2013, Zhang et al. 2021).
I tried to recreate the feeling of this common yet lovely type of scene in sci-fi movies where a ship or station gets dwarfed by a gigantic object slowly emerging behind it from the shadows - the only difference is that the 'giant' eye here is only about 3 cm wide, though that was still huge for the time.
References and technical details about the reconstruction under the cut:
The soft parts of I. glaessneri are not known (except for the eyes). Trunk appendages are based on I. curvirostratus (Zhang et al. 2021). Great appendages are partially based on I. communis, which may be the adult form of I. glaessneri (Fu et al. 2012); unfortunately, the great appendages of I. communis are poorly preserved (García-Bellido et al. 2009), so frontal appendage morphology was complemented with the better-known I. acutangulus.
The Isoxys is depicted here with only 11 pairs of trunk limbs, instead of the usual 13+ (Zhang et al. 2021). Based on the assumption that the ancestral arthropod grew by post-hatching addition of segments (anamorphosis) (Liu et al. 2016), a reduced number of trunk limbs was judged appropriate given the small size of the specimen (ca. 6.5 mm) and the possible juvenile nature of I. 'glaessneri'.
References:
Fu, D., Zhang, X., Budd, G. E., Liu, W., & Pan, X. (2014). Ontogeny and dimorphism of Isoxys auritus (Arthropoda) from the Early Cambrian Chengjiang biota, South China. Gondwana Research, 25(3), 975–982. https://doi.org/10.1016/j.gr.2013.06.007
García-Bellido, D. C., Paterson, J. R., Edgecombe, G. D., Jago, J. B., Gehling, J. G., & Lee, M. S. Y. (2009). The bivalved arthropods Isoxys and Tuzoia with soft-part preservation from the Lower Cambrian Emu Bay Shale Lagerstätte (Kangaroo Island, Australia). Palaeontology, 52(6), 1221–1241. https://doi.org/10.1111/j.1475-4983.2009.00914.x
Legg, D. A., & Vannier, J. (2013). The affinities of the cosmopolitan arthropod Isoxys and its implications for the origin of arthropods. Lethaia, 46(4), 540–550. https://doi.org/10.1111/let.12032
Liu, Y., Melzer, R., Haug, J., Haug, C., Briggs, D., Hörnig, M., He, Y., & Hou, X. (2016). Three-dimensionally preserved minute larva of a great-appendage arthropod from the early Cambrian Chengjiang biota. Proceedings of the National Academy of Sciences, 113, 5542–5546. https://doi.org/10.1073/pnas.1522899113
Paterson, J. R., Edgecombe, G. D., & García-Bellido, D. C. (2020). Disparate compound eyes of Cambrian radiodonts reveal their developmental growth mode and diverse visual ecology. Science Advances, 6(49), eabc6721. https://doi.org/10.1126/sciadv.abc6721
Schoenemann, B., & Clarkson, E. N. k. (2011). Eyes and vision in the Chengjiang arthropod Isoxys indicating adaptation to habitat. Lethaia, 44(2), 223–230. https://doi.org/10.1111/j.1502-3931.2010.00239.x
Zhang, C., Liu, Y., Ortega-Hernández, J., Wolfe, J. M., Jin, C., Mai, H., Hou, X. G., Guo, J., & Zhai, D. (2021). Differentiated appendages in Isoxys illuminate origin of arthropodization. Research Square.
#echidnacaris#anomalocaris#anomalocaris daleyae#taxa named after people i know ._.#radiodont#arthropod#emu bay shale#australia#cambrian#paleozoic
485 notes
·
View notes
Text

The orthid brachiopods Nanorthis bifurcata and Lipanorthis santalaurae (attached) from the Early Ordovician of Argentina.
Got inspired by a cool paper I read today and decided to try my hand at 2D art for once. I'm not very good at it but I have to admit it's pretty fun.
Reference:
Santos, A., Mayoral, E., Villas, E., Herrera, Z., & Ortega, G. (2014). First record of Podichnus in orthide brachiopods from the Lower Ordovician (Tremadocian) of NW Argentina and its relation to the early use of an ethological strategy. Palaeogeography, Palaeoclimatology, Palaeoecology, 399, 67–77. https://doi.org/10.1016/j.palaeo.2014.02.003
#nanorthis#lipanorthis#orthid#brachiopod#ordovician#paleozoic#paleontology#palaeoblr#paleoart#my art
39 notes
·
View notes
Text

Conulariids are an extinct group of probable cnidarians with 4-sided pyramidal thecae. They are relatively uncommon fossils but ranged from the Cambrian (possibly Ediacaran?) to the Triassic, comprising tens of genera and hundreds of described species (Lucas 2012).
Here are the reconstructed thecae of a small selection of species from every period of the conulariids' range, starting from the Cambrian in the top left and reaching all the way to the Triassic in the bottom right.
Since their soft parts are virtually never preserved (due to them being cnidarians and all that) (Van Iten & Südkamp 2010), most of our knowledge of conulariid biology and evolution is based on their more fossil-friendly thecae, which were composed of thin organophosphatic lamellae (Leme et al. 2008). Live conulariids were attached to the substrate by the apex of their theca; they probably captured suspended food particles or small prey using tentacles, just like other cnidarians, but it's hard to go in any more (non-speculative) detail without preserved soft tissues.
References:
Babcock, L. E. (1986). Devonian and Mississippian conulariids of North America. Part B. Paraconularia, Reticulaconularia, new genus, and organisms rejected from Conulariida. Annals of the Carnegie Museum, 55, 411–479. https://doi.org/10.5962/p.215204
Guimarães Simões, M., Coelho Rodrigues, S., Moraes Leme, J. de, & Van Iten, H. (2003). Some Middle Paleozoic Conulariids (Cnidaria) as Possible Examples of Taphonomic Artifacts. Journal of Taphonomy, 1(3), 163–184.
Hughes, N. C., Gunderson, G. O., & Weedon, M. J. (2000). Late Cambrian Conulariids from Wisconsin and Minnesota. Journal of Paleontology, 74(5), 828–838. https://doi.org/10.1666/0022-3360(2000)074<;0828:LCCFWA>2.0.CO;2
John, D. L., Hughes, N. C., Galaviz, M. I., Gunderson, G. O., & Meyer, R. (2010). Unusually preserved Metaconularia manni (Roy, 1935) from the Silurian of Iowa, and the systematics of the genus. Journal of Paleontology, 84(1), 1–31. https://doi.org/10.1666/09-025.1
Leme, J. M., Simões, M. G., Marques, A. C., & Van Iten, H. (2008). Cladistic Analysis of the Suborder Conulariina Miller and Gurley, 1896 (cnidaria, Scyphozoa; Vendian–Triassic). Palaeontology, 51(3), 649–662. https://doi.org/10.1111/j.1475-4983.2008.00775.x
Lucas, S. (2012). The Extinction of the Conulariids. Geosciences, 2, 1–10. https://doi.org/10.3390/geosciences2010001
Sendino, C., & Zagorsek, K. (2011). The Aperture and Its Closure in an Ordovician Conulariid. Acta Palaeontologica Polonica, 56, 659–663. https://doi.org/10.4202/app.2010.0028
Slater, I. L. (1907). A monograph of British Conulariæ. Printed for the Palæontographical Society.
Thomas, G. A. (1969). Notoconularia, a New Conularid Genus from the Permian of Eastern Australia. Journal of Paleontology, 43(5), 1283–1290.
Van Iten, H., Konate, M., & Moussa, Y. (2008). Conulariids of the Upper Talak Formation (Mississipian, Visean) of Northern Niger (West Africa). Journal of Paleontology, 82(1), 192–196. https://doi.org/10.1666/06-083.1
Van Iten, H., Muir, L., Simões, M. G., Leme, J. M., Marques, A. C., & Yoder, N. (2016). Palaeobiogeography, palaeoecology and evolution of Lower Ordovician conulariids and Sphenothallus (Medusozoa, Cnidaria), with emphasis on the Fezouata Shale of southeastern Morocco. Palaeogeography, Palaeoclimatology, Palaeoecology, 460, 170–178. https://doi.org/10.1016/j.palaeo.2016.03.008
Van Iten, H., & Südkamp, W. H. (2010). Exceptionally preserved conulariids and an edrioasteroid from the Hunsrück Slate (Lower Devonian, SW Germany). Palaeontology, 53(2), 403–414. https://doi.org/10.1111/j.1475-4983.2010.00942.x
Waterhouse, J. B. (1979). Permian and Triassic conulariid species from New Zealand. Journal of the Royal Society of New Zealand, 9(4), 475–489. https://doi.org/10.1080/03036758.1979.10421833
敏郎杉山. (1942). 156. 日本産Conularidaの研究. 日本古生物学會報告・紀事, 1942(25), 185-194_1. https://doi.org/10.14825/prpsj1935.1942.185
#conularia#archaeoconularia#eoconularia#pseudoconularia#metaconularia#paraconularia#notoconularia#conulariid#cnidarian#paleozoic#mesozoic#palaeoblr#paleoart#my art
38 notes
·
View notes
Text

Protomonaxonid sponges standing around (as sponges do) on the enthusiastically bioturbated sediment of the Fezouata Shale, 478 million years ago in the Antarctic Circle.
These early demosponges have been given the rather cumbersome designation of 'Hamptonia' christi Form B, pending a formal description. Standing a few centimeters tall, they (and several other sponges found in Fezouata) formed dense but single-species assemblages interpreted as rapid and repeated colonization events in a hostile environment (Botting 2016).
I could not resist giving them the same coloration as their fossils, which are beautifully rendered in hues of iron oxides. Surely reds and oranges aren't unusual colors for sea sponges?
References:
Botting, J. P. (2016). Diversity and ecology of sponges in the Early Ordovician Fezouata Biota, Morocco. Palaeogeography, Palaeoclimatology, Palaeoecology, 460, 75–86. https://doi.org/10.1016/j.palaeo.2016.05.018
Van Roy, P. (2006). Non-trilobite Arthropods from the Ordovician of Morocco. Gent University.
#hamptonia#protomonaxonid#sponge#fezouata#ordovician#paleozoic#paleontology#palaeoblr#paleoart#my art
107 notes
·
View notes
Text

Generic belemnite, surrounded by equally generic Aurelia-type jellyfish.
#mostly modelled from memory so probably not 100% accurate#belemnite#cephalopod#mollusk#aurelia#scyphozoa#cnidarian#jurassic#mesozoic#paleontology#palaeoblr#paleoart#my art
109 notes
·
View notes
Text

Vendia sokolovi, a 1 cm-long enigmatic* organism from the upper Ediacaran of Russia (displaying the 'glide symmetry' characteristic of many Ediacaran forms), feeding (?) on the bacterial mats that used to be widespread before the Cambrian explosion.
*as Ediacaran organisms tend to be, alas
Some art I made a long time ago but still find kind of neat.
References:
Ivantsov, A. (2001). Vendia and Other Precambrian 'Arthropods'. Paleontological Journal, 35(4), 335-343. https://www.academia.edu/2605872/Vendia_and_Other_Precambrian_Arthropods_
Ivantsov, A. (2004). New Proarticulata from the Vendian of the Arkhangel’sk Region. Paleontological Journal, 38(3), 247–253.
#vendia#proarticulata#ediacaran biota#whatever they are#ediacaran#proterozoic#precambrian#paleontology#palaeoblr#paleoart#my art
405 notes
·
View notes
Text

A shortfin mako shark (Isurus oxyrinchus) I made for an introductory course to ZBrush.
#could be better could be worse#shortfin mako shark#mako shark#shark#isurus#chondrichthyes#vertebrate#holocene#zbrush#my art
38 notes
·
View notes
Text
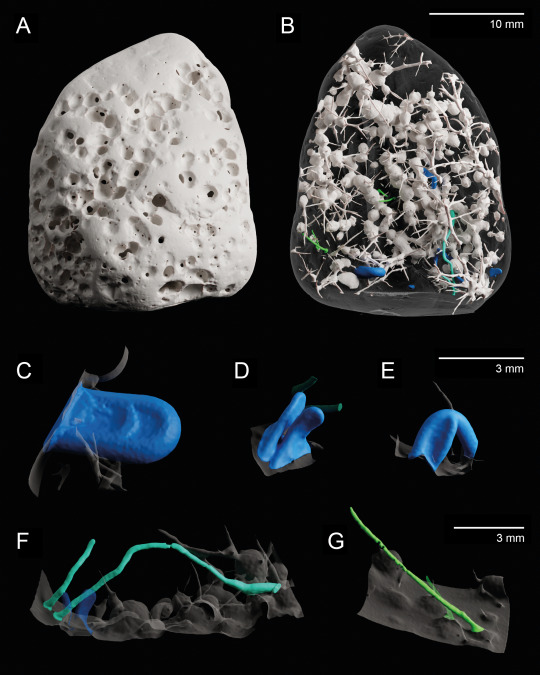
X-ray microtomography 3D model of a bioeroded limestone pebble given to me by a friend who wanted to know what the holes were. Not technically art but I think it looks cool.
I identified 4 distinct types of borings (ichnotaxa) in this 3-cm pebble:
Entobia cf. cretacea (white): typical sponge borings, composed of interconnected chambers and exploratory threads; by far the dominant ichnotaxon in the sample (97% of bioeroded volume).
Caulostrepsis taeniola (blue): U-shaped borings made by polychaete worms (e.g. spionids).
Unnamed Caulostrepsis ichnospecies (cyan): long and tightly U-shaped borings, possibly also made by polychaete worms.
Trypanites solitarius (green): simple unbranched borings that can be produced by many different organisms (polychaetes, bryozoans, phoronids).
(A-B) Full sample. (A) External view. (B) Virtual mould of the borings. (C-G) Virtual moulds of individual borings. (C-E) Caulostrepsis taeniola (note the variability in size). (F) Caulostrepsis unnamed ichnospecies. (G) Trypanites solitarius.
Experiments with limestone bricks show that bioerosion comparable in nature and extent to the one showcased here can develop after 3-4 years (Färber et al. 2016), providing a rough estimate of the time elapsed between this rock fragment first being exposed and it washing onto the shore where my friend picked it up.
References:
Bromley, R. G. (1972). On some ichnotaxa in hard substrates, with a redefinition of Trypanites Mägdefrau. Paläontologische Zeitschrift, 46(1), 93–98. https://doi.org/10.1007/BF02989555
Bromley, R. G. (2004). A Stratigraphy of Marine Bioerosion. Geological Society, London, Special Publications, 228, 455–479. https://doi.org/10.1144/GSL.SP.2004.228.01.20
Bromley, R. G., & D’Alessandro, A. (1983). Bioerosion in the Pleistocene of Southern Italy: Ichnogenera Caulostrepsis and Maeandropolydora. Rivista Italiana Di Paleontologia e Stratigrafia, 89(2), Article 2.
Bromley, R. G., & D’Alessandro, A. (1984). The ichnogenus Entobia from the Miocene, Pliocene and Pleistocene of southern Italy. Rivista Italiana Di Paleontologia e Stratigrafia, 90(2), Article 2.
Färber, C., Titschack, J., Schoenberg, C., Ehrig, K., Boos, K., Baum, D., Illerhaus, B., Asgaard, U., Bromley, R., Freiwald, A., & Wisshak, M. (2016). Long-term macrobioerosion in the Mediterranean Sea assessed by micro-computed tomography. Biogeosciences, 13, 3461–3474. https://doi.org/10.5194/bg-13-3461-2016
#entobia#caulostrepsis#trypanites#sponge#polychaete#bioerosion#trace fossil#holocene#ichnology#ichnotaxonomy#paleontology#geology#tomography#microtomography
110 notes
·
View notes
Text

Odaraia alata, (yet another) strange arthropod of the Burgess Shale.
Unlike many of the 'weird wonders' of the Burgess like Hallucigenia or Opabinia, Odaraia was quickly identified as an arthropod (although its precise position remains unclear) but what sets it apart is its completely unique functional morphology: in addition to a very unusual telson with three flukes (quite reminiscent of the tail of a plane), Odaraia had a bivalved carapace that almost completely enclosed its body (Briggs & Whittington, 1981), like a living burrito with many legs.
Since the legs were unable to reach out beyond the shield, Odaraia must have spent its life swimming (possibly on its back like a fairy shrimp, based on its center of gravity), using its legs to create currents to guide food particles into the shield space and towards the mouth; the tail flukes likely assisted in stabilizing and steering. The whole animal was about 15 cm long (Briggs & Whittington, 1981).
In the background are Pirania muricata sponges, contemporary to Odaraia and also found in the Burgess Shale (Walcott, 1924).
References and notes about the reconstruction:
The cephalic appendages of Odaraia are poorly known; this reconstruction is based on the interpretation of Briggs & Whittington (1981), which consists of a pair of denticulated, mandible-like structures and, anterior to it, two possible pairs of simple segmented appendages - but the latter are highly uncertain and should be taken with caution.
Briggs, D. E. G., & Whittington, H. B. (1981). The arthropod Odaraia alata Walcott, Middle Cambrian, Burgess Shale, British Columbia. Philosophical Transactions of the Royal Society of London. B, Biological Sciences, 291(1056), 541–582. https://doi.org/10.1098/rstb.1981.0007
Budd, G. E. (2008). Head Structure in Upper Stem-Group Euarthropods. Palaeontology, 51(3), 561–573. https://doi.org/10.1111/j.1475-4983.2008.00752.x
Charles D. Walcott. (1924). Middle Cambrian Spongiae. In Cambrian Geology and Paleontology (Vol. 67, pp. 261–364). Smithsonian Institution. https://www.biodiversitylibrary.org/item/9442
#odaraia#arthropod#pirania#protomonaxonid#sponge#burgess shale#cambrian#paleozoic#paleontology#palaeoblr#paleoart#my art
131 notes
·
View notes
Text

Epibiosis - An interesting ecological association as it might have looked 508 millions years ago: a cluster of Micromitra burgessensis, a species of early brachiopod and without doubt one of the prettiest organisms of the Cambrian, attached to the enigmatic tube-building animal Tubulella sp., a possible cnidarian.
In the Burgess Shale, almost a third of M. burgessensis individuals are preserved attached to another organism - often the spiny sponge Pirania, but they weren't too picky, settling on many different biogenic substrates (such as the tube here), and even other individuals of their own species! The strong tendency of Micromitra (and several other brachiopods species) in this deposit to colonize living organisms is most likely due to the rarity of other hard substrates like rocks, and a predominantly soft, soupy seafloor that did not provide suitable attachment points (Topper et al. 2015a).
Speculative hydroid-like polyps are also depicted on the tube, inspired by modern tube worms covered in hydroids that I saw while looking for reference pictures. Despite the exceptional range of soft bodied fossils collected in the Burgess Shale, no cnidarian soft tissues are known from this deposit, as the conditions required for their preservation appear different to those for other animals (Hancy & Antcliffe 2020): one wonders how many soft-bodied epibionts have disappeared without leaving a single trace in the fossil record, despite affecting the life of their hosts and possibly even competing with other, more fossil-friendly epibionts like brachiopods.
(Reconstruction of fossil specimen ROM63170 as figured in Topper et al. 2015b. Additional reconstruction information based on Topper et al. 2015a, 2015b. Maximum shell width of the center brachiopod is approx. 5 mm.)
References:
Hancy, A. D., & Antcliffe, J. B. (2020). Anoxia can increase the rate of decay for cnidarian tissue: Using Actinia equina to understand the early fossil record. Geobiology, 18(2), 167–184. https://doi.org/10.1111/gbi.12370
Topper, T. P., Strotz, L. C., Holmer, L. E., & Caron, J.-B. (2015). Survival on a soft seafloor: Life strategies of brachiopods from the Cambrian Burgess Shale. Earth-Science Reviews, 151, 266–287. https://doi.org/10.1016/j.earscirev.2015.10.015
Topper, T. P., Strotz, L. C., Holmer, L. E., Zhang, Z., Tait, N. N., & Caron, J.-B. (2015). Competition and mimicry: The curious case of chaetae in brachiopods from the middle Cambrian Burgess Shale. BMC Evolutionary Biology, 15(1), 42. https://doi.org/10.1186/s12862-015-0314-4
#micromitra#paterinid#brachiopod#tubulella#cnidarian#epibiont#symbiont#burgess shale#cambrian#paleozoic#paleontology#palaeoblr#paleoart#my art
77 notes
·
View notes